|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
2 Биология виноградного растенияВиноград — одно из древнейших цветковых растений земного шара. В далеком прошлом он произрастал на открытых, хорошо освещенных солнцем местах и имел вид кустарника с прямостоячими побегами, характеризующимися моноподиальным ростом. Примерно в третичном периоде, когда под влиянием теплого и влажного климата тропические леса стали занимать все большие свободные пространства, многие представители семейства Vitaceae оказались в тенистых лесах, где вынуждены были приспосабливаться к непривычным для них условиям обитания. Среди многочисленных видов лесного сообщества у растений винограда в процессе длительной эволюции вырабатывались особенности, позволяющие приспособиться к новым условиям обитания. Изменялись отдельные свойства и биологические функции органов в жизненном цикле развития, которые были закреплены генетически путем естественного отбора. Это и привело к изменению формы виноградного растения от кустовидной к лазящей древесной лиане, не имеющей прочного ствола и сучьев, как у других лесных древесных растений, с тонкими (не более 15 мм) и длинными (до 20 м) побегами, снабженными усиками. Верхушка побега преобразовалась в усик, моноподиальный рост стебля сменился симподиальным. С помощью усиков растение прочно цеплялось за деревья, побеги его быстро росли, взбирались вверх и выносили ассимиляционный аппарат к свету из — под полога леса. В случае повреждения стебля, поломки его ветром или в результате других механических воздействий и неблагоприятных факторов среды у виноградной лианы выработалась способность быстро восстанавливать рост за счет развития расположенных на побеге пазушных скороспелых пасынковых почек, а также центральной, замещающих почек глазка и спящих почек. Причем построение большой массы ассимилирующих органов — вегетирующих побегов и листьев — стало проходить у виноградного растения без развития мощного скелета (ствол, сучья, ветви). Ч. Дарвин писал, что лазящие растения удивительно мало затрачивают органического вещества для выноса растения к свету, воздуху и созданию обширной поверхности своей листвы для улавливания солнечных лучей. Это целиком можно отнести и к винограду. Быстрой подаче и продвижению воды с растворенными в ней питательными элементами по длинному и тонкому стеблю от корней к листьям способствовали большая (147 кПа — до 1,5 атм) сосущая сила корней, обусловленная высоким осмотическим давлением, и активная транспирация. В свою очередь, хорошо развитое жилкование листьев и особенности строения проводящих сосудов древесины, трахей и ситовидных трубок, их большой размер и длина обеспечивают высокую пропускную способность и быстрый отток продуктов ассимиляции, синтезированных в листьях, в побеги и корни. Поскольку надземная часть виноградного растения не имеет прочного скелета, может легко повреждаться и быть недолговечной, у него выработалось приспособительная способность накапливать запасы питательных веществ, главным образом в корнях — в тканях либриформа, лубяных волокнах и сердцевинных лучах. Запасные вещества расходуются растением в самые критические периоды жизни — при ежегодном весеннем распускании почек, росте и развитии побегов, листьев и репродуктивных органов, а также способствуют выживанию виноградного растения после суровых зим. На основе использования этой биологической особенности разработаны направленные агротехнические приемы по восстановлению кустов винограда, поврежденных морозами и заморозками (обновление плантажа с подрезкой корневой системы и др.). В стеблях питательных веществ накапливается значительно меньше, чем в корнях, и они концентрируются главным образом на узлах в диафрагме и расходуются растением также преимущественно весной на питание распускающихся почек и первоначальный рост побегов. Для удержания виноградного растения в пространстве с помощью усиков в процессе длительного исторического развития выработались особенности строения стебля, обеспечивающие его малую массу и прочность: членистые побеги, состоящие из узлов и междоузлий; отсутствие тяжелой твердой механической ткани (склеренхимы); большой размер и рыхлое строение всех тканей стебля; раннее пробкование паренхимных клеток сердцевины, вследствие чего междоузлия побега становятся полыми и заполняются воздухом. Это усиливает газообмен в растении, высыхание коры в период вызревания побегов и превращение ее в легкую корку. Отличительная биологическая особенность виноградного растения — определенная взаимосвязь между ростом и плодоношением. Так, например, если у ряда плодовых культур, в частности у малины, побеги в первый год растут, а во второй год дают урожай, у яблони для плодоношения имеются специальные образования — копьеца, плодушки и другие, то у винограда вегетативный рост и генеративные процессы сочетаются в одном органе — побеге, который может быть плодоносным и бесплодным, а листья того и другого побега обеспечивают гроздь вырабатываемыми в них ассимилятами. Виноградной лиане, произрастающей в естественных условиях, свойственно саморегулирование роста и плодоношения. Оно состоит в том, что из множества заложившихся на побегах почек весной в силу внешних и внутренних факторов большая часть их не развивается. Не развиваются в грозди и некоторые соцветия, и только незначительная часть цветков образует ягоды. Благодаря этому у виноградного растения отсутствует периодичность плодоношения. При культивировании винограда она также не наблюдается, поскольку применение различных агротехнических приемов (обломка, обрезка, нагрузка кустов глазками и побегами и др.) позволяет регулировать рост и плодоношение виноградного куста и тем самым ежегодно получать высокие урожаи хорошего качества. В отличие от других древесных растений у винограда более сильно выражена продольная и плоскостная полярность. По М. X. Чайлахяну, полярность является неотъемлемым свойством органической материи и выражается в морфофизиологических различиях на противоположных концах клеток, тканей, органов и всего растения. Она проявляется в полярно противоположных способностях— листостеблевой — на основе воздушно-светового питания и корнеобразовательной — на основе водно-почвенного питания. Борьба и единство этих способностей принимается как движущая сила индивидуального развития растений. Один из важных внутренних факторов растений, обусловливающих направленность протекания этих процессов и их активность, — деятельность фитогормонов (ауксины, гиббереллины, ингибиторы), которые образуются преимущественно в зонах с высокой меристематической активностью — в верхушке стеблей и корней, в молодых листьях, почках и растущих плодах. Все они — продукты метаболизма растений. Проникая в ткани, обладающие высокой чувствительностью, фитогормоны действуют на клетки — рецепторы, вызывая их активное деление и растяжение. Рост корней определяется преимущественно действием ауксинов с редуцированным содержанием ингибитора; рост побегов — в большей степени совместным действием гиббереллина и ингибитора; рост семян проходит при воздействии гиббереллина, цитокинина или ауксина с редуцированным содержанием эндогенного ингибитора. Фитогормоны поступают вначале к верхушечным почкам, а затем к нижним. В первую очередь и наиболее интенсивно развиваются самые удаленные от основания побега почки: из них развиваются наиболее сильные побеги. Нижерасположенные почки распускаются позже, а самые нижние, если и распускаются, то из них развиваются более слабые побеги. Интенсивность роста побегов винограда очень высокая — до 10 см в сутки. На нее в значительной степени влияет пространственное расположение побега — вертикальное, наклонное или горизонтальное. Вначале, когда побег растет вертикально, почти не развиваются побеги 2-, 3–го и последующих порядков ветвления (пасынки), способствующие развитию листовой поверхности и обладающие свойством восстанавливать корреляции между корнями и ассимиляционной поверхностью. Здесь проявляется ингибиторное (тормозящее) действие верхушки на боковые почки. В случае прекращения верхушечного роста резко стимулируется рост пасынков. Они активно растут и тогда, когда побег занимает горизонтальное или наклонное положение, то есть при подавлении продольной полярности. Это используют в виноградарстве, если необходимо вызвать рост боковых побегов — пасынков. Плоскостная полярность у винограда проявляется в дорзовентральном, асимметричном строении всех органов: побега, черешка листа, листовой пластинки, корней и семян. Асимметричность строения органов обусловлена значительными различиями их физиолого-биохимических свойств и разнообразием выполняемых функций. Дорзовентральность корней способствует лучшему продвижению их в почву. У растущих побегов винограда плоскостная полярность выражается в асимметричном строении. В поперечном сечении побег имеет вид прямоугольника с закругленными углами и неравномерно развитыми четырьмя сторонами — брюшной, спинной, желобчатой и плоской. Брюшная сторона широкая, более развитая, однако степень роста у нее слабее, чем у спинной стороны, поэтому верхушка растущего побега загибается книзу. Спинная сторона более выпуклая и интенсивней окрашена. Брюшной стороной побег стелется по земле или прикрепляется к опоре. При росте побега спинная и брюшная стороны сохраняют в пространстве свое положение по всей его длине. Желобчатая и плоская стороны чередуются от одного узла к другому. Этим объясняется размещение листьев на побеге в чередующемся порядке, что способствует наиболее полному использованию ими света. Такое размещение сторон побега в пространстве способствует лучшему ветвлению стебля и расположению пасынковых побегов 1-, 2–го и последующих порядков, что также является приспособительной биологической особенностью, свойственной лианам. Зная закономерности роста и развития побегов, можно направленно влиять на проявление свойств полярности путем соответствующей подвязки побегов в определенном направлении и под определенным углом, удаления части побегов, проведения обломки их в начале вегетации, а также путем пасынкования, прищипки, чеканки побегов. Ослаблять или усиливать проявление полярности — это значит активно вмешиваться в биологические свойства виноградного растения и тем самым направлять прохождение биологических процессов в нужном направлении. Важная биологическая особенность виноградного растения — наличие корреляции между ростом и развитием отдельных органов, в частности между развитием корневой системы и ростом надземной части куста, ростом и развитием вегетативных и репродуктивных органов. Изучение этой и других рассмотренных биологических особенностей у винограда, разработка путей и способов управления ими — теоретическая основа всех агротехнических приемов по системе ведения культуры и формированию кустов, их обрезке, установлению оптимальной нагрузки глазками, побегами и урожаем, подвязке побегов, комплексу операций с зелеными частями куста, применению регуляторов роста, технологий выращивания посадочного материала. Виноград в отличие от многих древесных культур — наиболее пластичное растение, и в промышленных насаждениях приходится работать с каждым сортом и кустом в отдельности. Для этого необходимо знать биологические особенности, строение органов виноградного растения, выполняемые ими функции. Строение, рост, развитие органов виноградного растения и их функцииК вегетативным органам виноградного растения относятся корень, стебель и листья, к генеративным (репродуктивным) — цветки, грозди, ягоды и семена. Почки у винограда могут осуществлять функции роста и плодоношения и давать начало вегетативным и одновременно генеративным органам. Корень и корневая система.Корни выполняют очень важную жизненную функцию растения. Они поглощают из почвы воду с растворенными в ней минеральными веществами, которые под давлением по проводящей системе передвигаются вверх в надземную часть к стеблям, листьям и генеративным органам. Молодые корни (мочки) наряду с растворами минеральных солей способны поглощать из почвы углекислый газ. Старые скелетные корни прочно закрепляют растение в почве. Они обычно длинные, мясистые, равномерно утолщенные, снаружи покрыты тонким слоем ежегодно отделяющейся корки. Скелетные корни служат не только проводниками воды с растворенными в ней питательными элементами по стеблю к листьям и генеративным органам. В них главным образом происходит синтез органических веществ, откладываются запасные питательные вещества (крахмал, белок, жиры), необходимые для растения в критические периоды его жизни. Исследованиями установлено, что при отсутствии пополнения запасов питательных веществ скелетные корни могут обеспечить жизнедеятельность куста до 6 лет. Совокупность молодых и старых корней составляет корневую систему растения. Важная функция корней, особенно молодых, — превращение углеводов в аминокислоты и осуществление первичного синтеза белка. Образовавшиеся в листьях в процессе фотосинтеза углеводы быстро передвигаются вниз по стеблю и в корнях превращаются в сахара и различные органические кислоты, которые, взаимодействуя с аммонийными солями, образуют смеси аминокислот — основных компонентов белковых веществ. Незначительная часть органических кислот выделяется корнями в почву и вместе с разложившимися остатками корней обогащает ее органическими веществами. Это способствует развитию и скоплению в прикорневом слое почвы полезных микроорганизмов (грибы, бактерии и др.) в результате жизнедеятельности которых труднодоступные для растений питательные элементы почвы превращаются в легкоусвояемые формы. Из корней основная часть аминокислот поднимается вверх и концентрируется в тканях растущего побега и гроздей, используется для построения вновь образующихся клеток. Формирование и рост молодых побегов и почек зависят от прохождения всех этих процессов. Корни винограда содержат 40–60% воды, по сравнению со стеблем в них больше азота, фосфора и зольных элементов, но меньше калия. Из пластических веществ в корнях больше всего углеводов, содержание которых заметно изменяется в течение сезона. В сентябре в корнях бывает 7% моносахаров, 3 — дисахаров и 13% крахмала. Наибольшее количество крахмала (23%) установлено в тонких корнях, наименьшее (15%) в толстых. Содержание крахмала увеличивается в ноябре. К. этому времени им заполнены коровая часть корня и сердцевинные лучи. В позднеосенний и зимний периоды до начала сокодвижения в корнях содержится 1,5–3% сахаров и свыше 20% крахмала. Кроме того, в зоне камбия корней имеются жиры, которые начинают накапливаться во второй половине лета (июле), а максимальное их количество наблюдается в зимний период. Характер развития корней и корневой системы в целом зависит от способа размножения винограда, видовых и сортовых особенностей, физико-механических свойств, химического состава почвы, ее температуры, влажности и метода предпосадочной обработки. При семенном размножении во время прорастания семени сначала через трещину его клювика из первичного корешка зародыша развивается главный, стержневой корень, на котором вскоре образуются корневые волоски. Через несколько дней главный корень начинает ветвиться, на его утолщениях появляются более тонкие боковые корни 1–го порядка, от которых отходят корни 2–го порядка, затем 3–го и т. д. От стержневого корня они идут сначала под прямым углом, а затем кончики их изгибаются и они растут почти параллельно главному корню. Стержневой корень сеянца растет сильнее боковых и в течение первого года может углубиться в почву до 1 м и более. В первые 2 года корневая система развивается значительно сильнее надземной части. У сеянца место перехода от корня к стеблю называют корневой шейкой. При размножении винограда вегетативным способом — черенками, прививкой, отводками — при благоприятных условиях для их укоренения на узлах, реже на междоузлиях, корни развиваются из корневых зачатков клеток перицикла стебля против сердцевинных лучей. В этом случае образуются придаточные корни, которые называют также адвентивными. Достигнув длины 8—10 см, они начинают ветвиться, образуя боковые корни разных порядков, имеющие на концах утолщения (мочки) белого цвета. При посадке коротких одноглазковых черенков, которые размещают обычно в горизонтальном положении, так же как и отводков, корни развиваются пучком с противоположной от глазка стороны. При посадке длинных 3-, 4-, 5–глазковых черенков, которые размещают вертикально, корни развиваются на узлах черенка, сначала на нижних, а затем и на верхних, но больше всего на базальной части черенка — пятке.
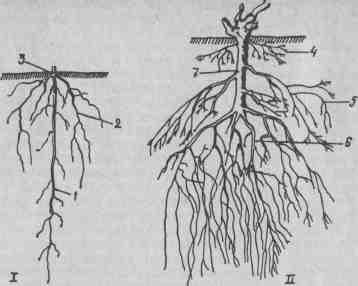
Рис. 16. Корневая система: / — сеянца, // — саженца; 1 — стержневой корень; 2 — боковые корни; 3— корневая шейка; 4 — поверхностные (росяные) корни; 5 — боковые корни; 6 — основные (главные) корни; 7 — подземный штамб. В отличие от сеянца корневая система саженца (растения, выращенного из черенка) имеет мочковатый характер. Подземная часть саженца состоит из стебля (подземный штамб) с отходящими от него несколькими ярусами корней (рис. 16). В верхней части подземного штамба на глубине 10–15 см в ежегодно обрабатываемом верхнем слое почвы развивается большое количество поверхностных (росяных) корней, или росособирателей. Эти корни тонкие, короткие, и если их своевременно не удалить проведением катаровки, то растение полностью перейдет на развитие корневой системы в поверхностном слое почвы, что препятствует развитию корней в глубоких слоях, приводит к ослаблению кустов и их полной гибели. Это наблюдается при продолжительной засухе летом и после сильных морозов зимой. На привитых виноградниках поверхностные корни удаляют особенно тщательно, в противном случае кусты перейдут на корни привоя, и цель прививки как одного из способов защиты винограда от филлоксеры не будет достигнута. В средней части подземного штамба, преимущественно на месте бывших узлов, развиваются боковые корни, а у самого его основания — главные (пяточные), наиболее толстые, длинные, глубоко проникающие в почву. К трем — четырем годам у виноградного куста развивается мощная корневая система. Корень растет путем деления клеток образовательной ткани — меристемы, находящейся в концевой его части в конусе нарастания. Как у стержневых, так и у адвентивных корней кончик корня размером в несколько миллиметров снаружи покрыт твердым, острым и очень прочным чехликом желтого цвета. У воздушных корней, которые легко развиваются в условиях повышенной температуры и влажности воздуха, чехлик коричневый. Под чехликом находится зона роста. Это наиболее нежная часть корня длиной 2–5 мм, белого цвета. Клетки ее, вытягиваясь в длину, толкают вперед чехлик, под прикрытием которого верхушка корня прокладывает себе путь между частицами почвы. Этим объясняется способность винограда расти на каменистых, скелетных и других твердых почвах. Наружные клетки чехлика постепенно высыхают, отслаиваются и взамен их на кончике корня непрерывно образуются новые клетки из конуса нарастания. За зоной роста идет зона поглощения. Она более утолщена, желто-белого цвета, длиной 2–7 см. Наружные клетки ее (эпидермис) вытягиваются в длину, образуя множество корневых волосков. Оболочка корневого волоска не имеет кутикулы, поэтому поглощаемая ими из почвы вода с растворами солей легко проникает в клетки. Корневые волоски недолговечны, живут 10–20 дней. По мере роста и удлинения корня старые волоски отмирают, а взамен их образуются новые, и зона поглощения перемещается. За зоной поглощения следует проводящая зона. Снаружи эта зона покрыта толстым слоем пробки коричневого цвета, с возрастом корней она превращается в корку. В месте перехода зоны поглощения в проводящую зону у корня заметна тонкая перемычка. До нее корень считается молодым, а после нее — скелетным. По анатомическому строению молодой корень (рис. 17) значительно отличается от старого, скелетного. На поперечном срезе молодого корня, сделанном в зоне поглощения, снаружи видны сомкнутые однородные клетки эпидермиса с корневыми волосками. Под эпидермисом находится первый слой коры — интеркутис. За ним следуют 20–25 слоев клеток коровой паренхимы, а затем слой клеток с уплотненными стенками — эндодерма. В середине корня находится центральный цилиндр, который у молодого корня состоит из двух — трех первичных сосудисто-волокнистых пучков, расположенных по кругу. К периферии от пучков находится несколько слоев паренхимных клеток, называемых перициклом, который дает начало боковым корням, поэтому его называют корнеродным слоем. К концу первого года вегетации у молодых корней в месте перехода поглощающей зоны в проводящую происходит ряд изменений. Корень покрывается пробкой буро-коричневого цвета, стареет, становится скелетным и приобретает вторичное анатомическое строение (рис. 18).
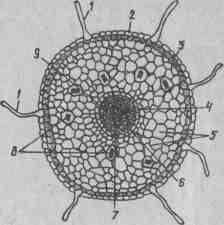
Рис 17. Анатомическое строение молодого корня в зоне поглощения (первичное строение): 1 — корневые волоски; 2 — эпидермис; 3 — интеркутис; 4 — эндодерма; 5 — первичная ксилема; б — первичная флоэма; 7 —кодовая паренхима; 8 — клетки, содержащие рафиды; 9 — перицикл.
Рис. 18. Анатомическое строение двулетнего корня (вторичное строение): / — кольцо древесины (ксилемы) первого года; 2 — кольцо древесины второго года; пд — перидерма второго года; ф— луб (флоэма); кам — камбий; ел — сердцевинные лучи; рл\ — радиальные лучи, заложившиеся весной первого года жизни корня; рл2—радиальные лучи, заложившиеся весной второго года; рл3 и рл — лучи, заложившиеся летом второго года; со1— сосуды древесины первого года; со2 и со3 — сосуды второго года (по Баранову). В центральном цилиндре между флоэмой и ксилемой и в промежутках между ними по кругу формируется образовательная камбильная ткань. Клетки камбия постоянно делятся и откладывают внутрь кольца вторичную ксилему (древесину), а наружу — вторичную флоэму (луб). Древесина и луб по радиусу пересекаются сердцевинными и радиальными лучами. В результате ежегодного образования камбия корень растет в толщину, и таким образом формируются годичные кольца древесины. Во вторичной флоэме камбий образует чередующиеся толстостенные и тонкостенные лубяные элементы твердого и мягкого луба. Наряду с этим клетки эндодермы пробковеют и вся первичная кора высыхает и постепенно слущивается. Из клеток перицикла впоследствии формируются кольца вторичной меристемы — феллоген (пробковый камбий). Снаружи все его ткани высыхают, образуя корку. Ежегодно в толще вторичной корки появляются новые кольца феллогена и новые слои корки. Корни разных видов винограда различаются по внешнему виду и анатомическому строению. У филлоксероустойчивых сортов американских видов, в частности у V. vulpina (V. riparia), корни более тонкие, сильно растут в длину, характеризуются компактным строением тканей, более мелкими сосудами проводящей системы, слаборазвитыми клетками паренхимы и меньшим их количеством, большей толщиной клеточных оболочек и клеток пробковой ткани. Интеркутис у них двухслойный, что считается одним из показателей устойчивости американских видов к филлоксере. Как свидетельствуют результаты исследований советских и зарубежных ученых, корни этих видов способны быстро изолировать поврежденные участки защитным слоем пробковой ткани, тогда как европейские сорта винограда этой способностью не обладают: на поврежденных филлоксерой участках изолирующий слой пробки у них образуется слабо, раны плохо заживают и загнивают. Большая часть боковых ответвлений корней американских видов отходит под углом 80°, или почти вертикально. У V. vinifera корни мясистые, толстые, рыхлого строения, с крупными сосудами ксилемы и боковыми ответвлениями, отходящими под небольшим углом. Корни не имеют периода покоя. Интенсивность их образования, роста и развития зависит от условий, в которых находится корневая система растения. При достаточном количестве тепла, влаги и питательных элементов в почве корни могут расти круглый год, образуя мочки как на молодых корнях, так и на скелетных. Однако в развитии корней ежегодно наблюдаются две наиболее интенсивные волны роста — весной, когда почва на глубине залегания основной их массы прогреется выше 6–8°C, и осенью, когда приостанавливается рост побегов, обильно выпадают осадки, а почва еще достаточно теплая (выше 7–8°C). Если же в период вегетации наступает продолжительная засуха, то второй волны роста не бывает. Активная жизнедеятельность и рост корней начинаются весной, до распускания почек, при температуре почвы 5–7°C, когда у растений происходит интенсивное сокодвижение («плач»). Жизнедеятельность корней усиливается в период интенсивного роста побегов и нарастания листового аппарата растения при температуре 18–24°C. Максимальный рост корней (до 12 мм в сутки) и их ветвление наблюдаются в первой половине лета, до фазы цветения, при оптимальной температуре почвы в корнеобитаемом слое 28–30°C. При более высокой температуре (34–40°C) наступает ослабление роста, повреждение корней и их гибель. В засушливый период года рост корней ослабевает. При снижении температуры почвы ниже 7–8СС их рост прекращается. Осенью корни покрываются коркой и пробковеют до чехлика, часть молодых неопробковевших корней отмирает, наступает массовый корнепад. Остаются только те корни, которые находятся в наиболее благоприятных условиях. Корнепад наблюдается круглый год, но наиболее активно он происходит в засушливый период года и зимой. Более долговечны, выносливы к засухе и низким температурам старые скелетные корни. Зимой в корнях образуются и накапливаются питательные вещества, в основном крахмал, сахара, жиры, которые расходуются растением весной следующего года как основной источник питания при распускании почек, первоначальном росте побегов, дифференциации плодовых образований и формировании урожая. В случае промерзания корнеобитаемого слоя почвы, по данным А. Г. Мишуренко, корни европейско-азиатских сортов винограда погибают при температуре —5–8°C, а амурского винограда и филлоксероустойчивых сортов подвоев американских видов — при —10–12°C. При благоприятных условиях зимовки весной рост корней возобновляется. Основная их масса образуется в первые 2 года после посадки. Количество основных корней и разветвлений первого порядка достигает в это время 60–70%. С возрастом растений число корней первого порядка уменьшается до 5–6 за счет отмирания самых тонких, но возрастает количество питающих корней, которые формируются главным образом на разветвлениях 3–4–го порядка. Эти корни долго остаются белыми. Основная масса корней на 2–й год размещается на глубине 20–80 см от поверхности почвы, на 3–й — на 20—130 и на 4–й год — 20—180 см. Глубина их размещения зависит от генетического типа почвы, ее физико-механических свойств, глубины предпосадочной обработки, условий выращивания, орошения. На плодородных, достаточно увлажненных легких и хорошо аэрируемых почвах корни мощные, глубоко проникают в почву и быстро ветвятся, образуя большое количество мелких корней. В оподзоленном черноземе и серой лесной почве с худшими физико-механическими свойствами они располагаются неглубоко. Так, на богатом питательными элементами карбонатном черноземе корни проникают до глубины 6 м, на карбонатном черноземе Молдавии 45% корней размещается на глубине 3–5 м, на каштановой почве и легком лёссовидном суглинке Таманского полуострова они проникают на глубину 3,5–4 м, на южном черноземе Крыма до 2,8, на легкой перегнойно-карбонатной почве Краснодарского края до 1,5, на серой лесной почве до 1,6, на торфяной почве Южного берега Крыма до 6 м. В южных засушливых районах на малоплодородных почвах с глубоким залеганием грунтовых вод корни слабо ветвятся, но проникают на глубину 5–6 м и более. Однако основная масса всасывающих корней растения располагается на глубине от 20–40 до 60 см (горизонт плантажной обработки почвы), где создаются наилучшие условия питания, аэрации, тепла и влажности. На почвах с близким (до 1,5 м) залеганием грунтовых вод корни виноградного растения размещаются в поверхностном слое над грунтовыми водами. В более северных районах виноградарства корневая система развивается в прогреваемом слое почвы на глубине 30–35 см. В горизонтальном направлении корни распределяются на расстояние от 2–5 до 8 м от куста, иногда заходят за пределы отведенного им пространства, особенно если нет растения — конкурента. Глубокая обработка междурядий способствует развитию корней в более глубоких слоях. На террасах корни лучше всего развиваются в средней полосе полотна, несколько слабее — в зоне выемки и наиболее слабо — на расстоянии 100 см от откоса террасы, что объясняется сильным просыханием почвы. Корни виноградного растения обладают хорошей регенеративной способностью. Они легко восстанавливаются при обрыве. Особенно интенсивно корни регенерируют при достаточном увлажнении почвы и внесении удобрений, образуя густую сеть мелких разветвлений. На способности хорошей регенерации корней основан прием обновления плантажа путем глубокого рыхления междурядий на винограднике. Если рост основных корней винограда задержать, то они начинают больше ветвиться, утолщаются и становятся более мощными. Чем сильнее развита корневая система растения, тем лучше рост и развитие всех органов надземной части куста и выше величина урожая. У сильнорослых сортов винограда, занимающих большую площадь питания, развивается и более мощная корневая система. Корневую систему виноградного растения в полевых условиях изучают путем раскопки. Зная глубину залегания и распространения основной массы питающих корней и применяя в промышленных насаждениях винограда соответствующую обработку почвы, внесение удобрений, орошение, защиту корней от низких температур и другие агротехнические приемы, позволяющие создавать наиболее благоприятные условия водно-воздушного режима и питания, необходимые для роста и развития корневой системы, можно получать ежегодно высокие урожаи хорошего качества. Стебель (ствол).Один из важнейших органов надземной части виноградного растения. По стеблю от корней в листья и в другие органы подается вода с растворенными в ней минеральными солями, а из листьев по стеблю поступают в корни растворы органических соединений — продукты фотосинтеза. В стеблях откладываются запасы питательных веществ, которые расходуются растением в критические периоды жизни — при распускании почек, росте молодых побегов и дифференциации соцветий весной будущего года. В отличие от древесных и кустарниковых растений стебель винограда не имеет осевой симметрии и четко выраженной постоянной Формы. У дикого винограда, произрастающего в условиях лесного сообщества, стебель имеет вид длинного оголенного гибкого и быстрорастущего ствола, высоко взбирающегося вверх по деревьям. В равнинных местах он свободно стелется по земле, занимая большие пространства. При выращивании в культуре виноградному растению придают вид куста со стеблем определенной формы, прямостоячего или наклонного, сильно разросшейся головы и отходящих от него многолетних ответвлений — рукавов, несущих на себе плодовые побеги с развивающимися на них вегетативными и репродуктивными органами. У растений, выращенных из семян, стебель начинается от корневой шейки, у выращенных из черенков — от основания (пятки) подземного штамба. С возрастом куста стебель утолщается и достигает 8—12 см и более в диаметре. Известны кусты винограда и с очень толстым стеблем. Так, в Калифорнии (США) есть 175–летний куст винограда со штамбом 210 см в окружности, занимающий 0,5 га. Надземная часть его поддерживается 60 столбами. Ежегодная урожайность куста составляет 10 т. На многолетних частях стебля, составляющих скелет куста, развиваются одногодичные плодовые побеги. Они членистые, состоят из узлов и междоузлий. На узлах формируются листья, соцветия, преобразующиеся впоследствии в грозди, а также пасынки, усики и пазушные почки. На междоузлиях побегов никакие органы не развиваются (рис. 19). Внутри каждого узла плодового побега имеется перегородка — диафрагма, состоящая из богатых крахмалом паренхимных клеток с толстыми одревесневшими оболочками. Живые ткани диафрагмы отделяются от мертвых клеток сердцевины несколькими слоями опробковевших клеток. Диафрагма — кладовая запасов питательных веществ стебля растения. На узлах побега, несущего соцветия или усики, диафрагма сплошная, в ней больше запасов питательных веществ; на узлах без усиков диафрагма неполная. Стебель растет как за счет деления клеток верхушечной меристемы конуса нарастания (апикальный верхушечный рост), так и за счет растяжения клеток междоузлиев (интеркалярный, вставочный рост). Однако деление клеток верхушечной меристемы на конце побега обеспечивает незначительное удлинение побега. Рост происходит главным образом за счет растяжения клеток. У основания побега первые 2–3 междоузлия растут недолго, поэтому остаются на побеге укороченными, следующие междоузлия нормальной длины. Максимальной длины достигает 5–6–е междоузлие. Междоузлия, несущие на узлах усики, растут наиболее интенсивно. Одновременно с ростом в длину стебель растет в толщину у самой верхней части он утолщается за счет увеличения размера клеток, а ниже — в результате образования вторичной меристемы (камбия), способствующего образованию концентрических слоев флоэмы и ксилемы. Стебель растет в толщину быстрее, чем корень.
Рис. 19. Плодоносный побег винограда: / — узел; 2 — междоузлие; 3 — листья; 4 — пазушная почка, 5 — пазушный побег (пасынок); 6 — усик; 7 — соцветия; 8 — верхушка побега (коронка); 9 — прошлогодний побег (лоза). Ветвится стебель за счет экзогенного заложения в конусе нарастания скороспелых пазушных почек, из которых в том же году развиваются пазушные побеги — пасынки. Они могут быть 1–го, 2–го и последующих порядков. В зависимости от экологических и погодных условий года, а также при определенных приемах агротехники и усиленном питании растений они могут развиться мощными и в тот же год дать урожай. Соцветия на пасынках формируются ускоренно. Иногда для усиления формирования кустов, получения дополнительного урожая и для других целей искусственно вызывают развитие пасынков путем прищипывания верхушки основного побега в ранние сроки. На одном побеге может развиться до 5–6 пасынков. Рост пасынковых побегов и их вызревание проходят быстрее, чем у основных побегов, несмотря на то, что пасынки развиваются позже. Наиболее сильно пасынки развиваются на нижних узлах побега, к вершине их рост постепенно затухает. При поломке или при повреждении верхушки основного побега его рост продолжает верхний пасынок. В отличие от основного побега пасынки менее развиты и у их основания только 1 недоразвитый листочек в виде чешуйки, в то время как у главного побега — 2. Усики на пасынках развиваются со 2–го, а на основном побеге — с 3–5 го узла. В пазухе листа пасынка закладывается зимующая почка (глазок), из которой на следующий год развивается основной побег, а на нем вновь пасынок и т. д. Самый интенсивный рост побегов совпадает с цветением, однако оптимальные температуры для роста побегов и цветения не совпадают. На интенсивность роста побегов сильно влияет температурный фактор. В начальный период роста, когда среднесуточные температуры воздуха еще низкие, побеги более энергично растут ночью. С повышением температуры различия в ночном и дневном приросте сглаживаются. На интенсивность роста побегов влияют полярность, сортовые особенности, экологические условия. При вертикальном положении стебля в рост трогаются сначала почки глазков верхней зоны, затем средней и нижней. В данном случае вышерасположенные побеги растут более интенсивно, чем нижерасположенные. При горизонтальном положении стебля распускание почек и рост побегов протекают более или менее равномерно. Это используют в практике виноградарства при подвязке рукавов и плодовых побегов в горизонтальном направлении. Сорта винограда обладают разной силой роста. Так, сорта восточной эколого-географической группы характеризуются большей силой роста, чем западноевропейской или группы бассейна Черного моря. Однако и внутри каждой группы есть сортовые различия. Каберне Совиньон более рослый, чем Алиготе и сорта группы Пино, а последние сильнее, чем сорта группы Шасла, Жемчуг Саба и др. До вступления виноградного растения в плодоношение все побеги у него выполняют функции вегетативных органов. С вступлением в плодоношение в одном побеге сочетается функция роста и плодоношения, что является характерной биологической особенностью винограда. Однако часть побегов остаются бесплодными, что зависит от генотипа сорта, внутренних и внешних условий в период закладки и формирования почек и зачатков соцветий в них. При обильных поливах, избыточном азотном удобрении, затенении и загущении кустов ростовые процессы преобладают над генеративными, и растение начинает жировать. В этом случае закладка плодоносных почек прекращается, и из почек развиваются бесплодные побеги. Иногда и из заложившихся плодоносных почек также развиваются бесплодные побеги. Не закладываются плодоносные почки и в случае очень слабого роста побегов. При гармоничном сочетании роста и плодоношения создаются наиболее благоприятные условия для закладки плодоносных почек и развития из них плодоносных побегов. Первоначально между 2–5–м узлом главная ось побега растет только верхушечной точкой роста. Это так называемый моноподиальный рост. Начинай с 3–6–го узла моноподиальный рост сменяется симподиальным. В этот период роста основная ось побега смещается и уступает место новому бугорку, который продолжает рост до следующего узла, где опять новая точка роста смещается и образуется новый бугорок, продолжающий рост побега. Смещенные точки роста образуют соцветие или усик. Такой рост побега путем смещения точки роста и замены ее новой точкой роста называют симподиальным. На 3–м узле точка роста снова продолжает расти вверх и усик или соцветие не образуется, то есть снова происходит моноподиальный рост, и т. д. Следовательно, соцветие или усик — это редуцированная верхушка побега, смещенного в сторону. Смена моноподиального роста симподиальным определяет закономерное чередование узлов на побеге, несущих на себе соцветия или усики, или не несущие их. В зависимости от расположения усика или соцветия по длине побега чередуется сторона его, несущая на себе пасынки и почки, со стороной, несущей усики и соцветия. При выращивании винограда из семян (у сеянца) стебель растет из стеблевой почечки, находящейся между двумя семядолями зародыша семени. С ростом подсемядольного колена (гипокотиля) зародышевая почечка выносится на поверхность почвы вместе с семядольными листьями. Стеблевая почечка зародыша, имеющая в конце нарастания форму бугорка, постоянно делится, и стебель растет в длину. Примерно через месяц после прорастания семени сбоку бугорка конуса нарастания из наружных клеток образуется другой бугорок — зачаток первого настоящего листа, и начинается дальнейший рост стеблевой части (эпикотиля). Затем через каждые 10–15 дней на каждой из противоположных сторон стебля вновь образуется по бугорку, которые дают начало новым листьям. У сеянцев листья закладываются по стеблю спирально. В пазухе бугорков листьев образуются новые бугорки — зачатки пазушных почек. Примерно до 6—10–го узла стебель сеянца растет моноподиально, то есть на узлах образуются только листья. Выше этих узлов наблюдаются некоторые изменения в росте стебля сеянца. Бугорок, образовавшийся в пазухе одного из листовых бугорков, растет быстрее обычного и вскоре сдвигает в сторону вершину конуса роста основного побега и замещает ее. Сдвинувшаяся в сторону вершина конуса роста в дальнейшем образует усик. Моноподиальный рост и ветвление сменяются симподиальным. Новая вершина побега растет, образуя новый листовой бугорок, в пазухе которого снова закладывается быстро развивающийся пазушный бугорок, сдвигающий основной конус роста. Так закладывается второй усик. На следующем узле смещение бугорка не наблюдается, и рост идет моноподиально. Формирование бугорков и отдельных органов у сеянца проходит медленно. Когда развивается первый настоящий лист, на вершине конуса нарастания закладывается уже бугорок 10–12–го узла. Рост стебля сеянца продолжается до конца периода вегетации. С наступлением периода покоя верхушечная почка отмирает. Со второго года вегетации побеги у сеянца развиваются из зимующих пазушных почек (глазков) и стебель растет не из верхушки побега, а симподиально. Вегетирующие побеги зеленые, травянистые, хрупкие, с изгибающейся вниз верхушкой. На узлах растущих побегов плодоносящих кустов в пределах 3—10–го узла от его основания, а иногда и выше развиваются соцветия или усики. На одном побеге может быть 1–2 соцветия и более. Побеги с соцветиями называют плодоносными, без соцветий — бесплодными; развившиеся на подземном штамбе — порослевыми, а на штамбе, рукавах, голове — волчками, или жирующими. Волчки и порослевые побеги обычно более толстые, отличаются сильным ростом, более рыхлым строением тканей и в год развития обычно не плодоносят. Заложившиеся на них зимующие почки в следующем году могут дать урожай. Волчки и порослевые побеги используют для формирования новых рукавов и омоложения кустов. Нередко в росте и развитии как основных, так и пасынковых побегов можно наблюдать различные аномалии в виде фасциации, то есть побег делается плоским и широким. В некоторых случаях образуется двойная верхушка: побег как бы состоит из двух сросшихся. К концу вегетации побеги замедляют рост и начинается их вызревание. Первые 3–7 междоузлий вызревают медленно, в течение 10—15 дней, далее темп вызревания резко возрастает. Вышерасположенные междоузлия (8— 14–е) вызревают в течение трех дней. Вызревание побегов — процесс анатомической и физиологической перестройки, наступающий после закладки перидермы. В этот период в клетках стебля откладываются запасы питательных веществ, возрастает содержание крахмала, уменьшается количество воды. У вызревших побегов к концу вегетации образуется кольцо пробкового камбия (феллоген), состоящее из нескольких слоев пробки, которая отделяет первичную кору от вторичной флоэмы. После этого кора начинает высыхать, образуя корку. У винограда она отделяется длинными полосами. Вследствие образования пробкового камбия и корки побеги в зависимости от сорта становятся светло-коричневыми до бурых и темно-коричневых. Хорошо вызревшие побеги называют лозой.
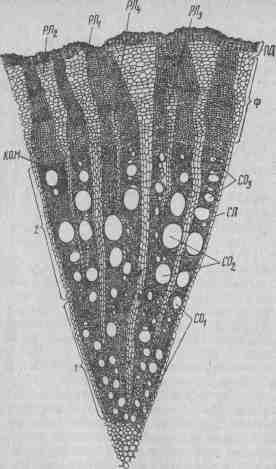
Рис. 20. Вызревший побег в поперечном разрезе: / — спинная сторона; 2 — брюшная; 3 — желобчатая; 4 — плоская; 5 — зимующая почка (глазок); 6 — усик. У вызревших однолетних побегов резко очерчивается округлая форма с явно выраженной асимметричностью сторон — дорзовентральностью (рис. 20). Самая широкая сторона побега брюшная. На ней наиболее развиты коленхима, перицикловые волокна, луб, древесина, сердцевинные лучи и др. Против брюшной стороны находится спинная сторона. Она более выпуклая и темно окрашена. На ней слабее развиты элементы коленхимы, древесины и другие, однако ростовые процессы здесь идут значительно активнее, чем на брюшной стороне. Менее развиты боковые стороны — желобчатая и особенно плоская. На желобчатой стороне расположен лист, а в его пазухе — пасынок и зимующая почка (глазок). Пасынок обращен к спинной, глазок — к брюшной стороне. На плоской стороне находится усик или гроздь. Анатомическое строение стебля (по Баранову). Вегетирующий побег в верхней части имеет первичное анатомическое строение, во многом сходное с анатомическим строением молодого корня. Наружные клетки первичной верхушечной меристемы конуса нарастания дифференцируются и образуют наружную кожицу— эпидермис, а внутренние слои — паренхимные клетки первичной коры: эндодермы и центрального цилиндра (рис. 21). Молодой побег отличается от корня меньшей толщиной первичной коры (8—10 слоев паренхимных более плотных клеток, богатых крахмалом), более развитой сердцевиной, эпидермисом с устьицами, выростами паутинистых волосков, щетинок и «жемчужных» железок. В первичной коре образуется коленхима. Центральный цилиндр состоит из пучков первичной флоэмы и ксилемы, расположенных радиально. Над ними находятся склеренхимные тяжи, которые называют волокнами перицикла, или перикамбиальными тяжами. Существуют некоторые различия в анатомическом строении узла и междоузлия. У последнего сильнее развиты паренхима и коленхима, слабее сосудистая система и больше диаметр сердцевины.
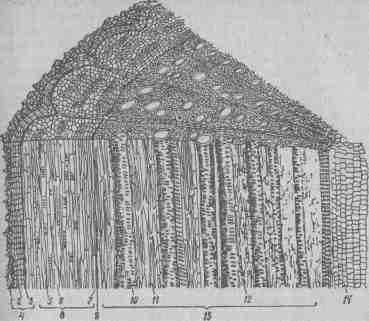
Рис. 21. Анатомическое строение стебля в конце первого года вегетации: / — корка; 2 — пробка; 3 — пробковый камбий (феллоген); 4— перидерма; 5 —лубяные волокна; 6 — ситовидные трубки; 7 —лубяная паренхима; 8 — луб (флоэма); 9 — камбий; 10 —1 пористые сосуды; 11—12 — перегородчатый либриформ; 13 — древесина (ксилема); 14 — сердцевина (по Баранову). Вторичное анатомическое строение стебля появляется после образования камбия между первичной флоэмой и ксилемой. В результате деятельности камбия побег растет в толщину. При этом образуется. вторичная флоэма с характерным слоистым строением, когда чередуются несколько слоев клеток мягкого и твердого луба, состоящего из ситовидных трубок — клеток — спутников, лубяных волокон и лубяной паренхимы. Стенки клеток твердого луба сильно утолщены. Волокна твердого луба выполняют роль механической ткани, которая придает прочность тонкостенной ткани мягкого луба. Вторичная ксилема состоит из сосудов — трахей и трахеид, механической ткани — перегородчатого либриформа и древесной паренхимы. Сердцевинные лучи у стебля имеют меньшую ширину, чем у корня, и служат для запаса пластических веществ, главным образом крахмала. В центре стебля находится сердцевина, состоящая из крупных пятиугольной формы омертвевших клеток. Лист.Главнейший орган виноградного растения, выполняющий основную физиологическую функцию — фотосинтез (способность синтезировать органические вещества из неорганических при помощи солнечной энергии, углекислого газа и воды). От работы листьев и их фотосинтетической активности зависит синтез углеводов, сахаров, аминокислот и других органических соединений, необходимых для нарастания вегетативной массы куста, формирования урожая и накопления сахара в ягодах. К. А. Тимирязев в работе «Жизнь растения» (1878) указывает, что в жизни листа выражается самая сущность растительной жизни, а растение — это лист. Лист у винограда простой, состоит из длинного черешка и листовой пластинки. У разных видов и сортов листья различаются по величине, форме, рассеченности, опушенности нижней стороны пластинки, ее изогнутости, а также по форме боковых вырезок, черешковой выемки, зубчиков, окаймляющих листовую пластинку, ее окраске и др. Сочетание морфологических признаков листа — характерный признак, который широко используют при определении: сортов. По черешку в листовую пластинку проходят сосудисто-волокнистые пучки, образующие 5 основных жилок. Разветвляясь, жилки формируют тонкую густую сетку, пронизывающую всю пластинку листа. По жилкам в лист поступает вода с минеральными солями и отводятся вырабатываемые листьями продукты ассимиляции. Развитие и рост листа начинаются с вытягивания бугорка растущей верхушки побега в удлиненную зачаточную пластинку. Молодые листочки защищены от неблагоприятных условий среды двумя пленчатыми прилистниками, которые после развития листьев опадают. Пластинка листа растет своим основанием в среднем 4–5 недель и имеет 3 волны роста. По данным К. Стоева и А. Амирджанова, в течение первых 6–9 дней пластинка листа растет медленно (2–8 см2 в сутки). Максимальный ее рост наблюдается во 2–ю и 3–ю недели (по 14–20 см2 в сутки). К концу 4–5–й недель интенсивность среднесуточного прироста пластинки снижается до 3–5 см2. Достигнув 30% своей величины, листья уже способны отдавать другим органам часть выработанных ассимилятов. Ко времени цветения более 50% листьев достигает максимальной величины, что значительно увеличивает продуктивность листовой поверхности побега и куста в целом. К периоду полного цветения 12 листьев из 19 становятся наиболее продуктивными. Ко времени завершения своего роста площадь листовой пластинки достигает оптимальной величины: в зависимости от сорта 120–300 см2 и более. Самые крупные листья находятся в пределах 6—12–го узла, самые мелкие — 21–25–го, промежуточную величину имеют листья нижнего яруса, в зоне 1–5–го узла. По длине побега листья у винограда располагаются в супротивно-чередующемся порядке, благодаря чему они не затеняют друг друга. С ростом побегов в длину увеличивается количество листьев на побеге, их размер, облиственность побега и всего куста. Оптимальная температура для роста листьев 28–30°C. При ее снижении, так же как и при повышении, интенсивность роста листьев ослабевает. На интенсивность роста листьев привитых растений существенно влияет корневая система подвоя. В частности, на подвое Рупестрис дю Ло листья сорта — привоя растут более интенсивно, чем у корнесобственных растений. Величина листовой поверхности зависит от силы роста побегов в длину, формы и нагрузки кустов побегами, густоты посадки, развития пасынков и др. По данным советских ученых, наибольшая поверхность продуктивных листьев (до 12 м2 на куст) развивается при формах куста типа одноплечего и двуплечего Гюйо. При малой чашевидной форме она составляет 8–9 м2. В условиях Средней Азии при больших веерных формах на двухплоскостной шпалере на 1 га насаждений развивается до 23 тыс. м2 листовой поверхности, на шпалере с козырьком— 21,8, вертикальной—16, при культуре врасстил — 8 тыс. м2. С повышением нагрузки средний размер листьев уменьшается, они становятся более мелкими. Густота посадки также влияет на величину листовой поверхности. При более густой посадке на 1 м2 площади питания приходятся 3,26, при редкой — 2,55 м2 листовой поверхности. В формировании последней принимают участие листья не только основных побегов, но и пасынков. Если пасынки не прищипывать, то на их долю приходится 40–50% общей листовой поверхности куста, а в случае их прищипывания на 4–5 узлов — 30–40%. Полное удаление пасынков уменьшает листовую поверхность куста на 30–55%. В загущенных посадках процесс фотосинтеза в затененных листьях и побегах прекращается. В пазухах таких листьев плодоносные почки не закладываются, а для своего роста и развития они вынуждены расходовать питательные вещества из других листьев. Поэтому очень важно, чтобы на винограднике все побеги с листьями были одинаково хорошо освещены. Процесс фотосинтеза наиболее интенсивно протекает на свету при температуре воздуха 28–30°C. В случае снижения ее фотосинтез ослабляется, а при температуре ниже 6°C прекращается. В процессе фотосинтеза большое значение имеет нормальное состояние хлоропластов и быстрый отток ассимилятов. У пожелтевших листьев, что наблюдается при их затенении, хлорозе и старении, деятельность пластид снижается. Результаты исследований советских и зарубежных физиологов показали, что в период вегетации фотосинтетическая активность листьев, расположенных на разных ярусах побега, неодинакова. В начале вегетации процесс фотосинтеза наиболее активно проходит в листьях нижнего яруса (от 5–го до 12–го узла побега), в середине и конце вегетации — в листьях среднего (от 15–го до 20–го узла) и верхнего ярусов. Отток ассимилятов из листьев начинается в момент, когда листья достигнут половины оптимальной величины. Молодые (2–3–недельные) листья не поставляют продуктов фотосинтеза, и рост побегов в этот период происходит за счет пластических веществ, ранее накопленных в других органах, — корнях и многолетних частях куста (рис. 22). Листья плодоносных побегов характеризуются большей ассимиляционной активностью, нежели бесплодных. Чем больше на кусте хорошо развитых листьев и чем интенсивнее их ассимиляционная активность, тем больше они вырабатывают углеводов и тем выше урожай винограда и его качество. Ассимиляционная поверхность листьев 28 тыс. м2 на 1 га виноградных насаждений наиболее продуктивна. В листьях происходят процессы не только ассимиляции, но и дыхания и транспирации, которые осуществляются при помощи многочисленных (140–190 на 1 мм2) устьиц, находящихся на нижней поверхности листа. При дыхании наблюдается окислительный распад сложных органических соединений (углеводов) до углекислоты и воды с выделением свободной энергии, обеспечивающей жизнедеятельность растения. Оптимальная температура для дыхания растения несколько выше, чем для фотосинтеза. Наиболее интенсивно оно проходит при температуре 30–40°C, в результате чего наблюдается значительная потеря углеводов. Большое значение при дыхании имеет набухание коллоидов плазмы, то есть насыщенность их водой. Для нормального дыхания, кроме соответствующей температуры и влажности, необходимы кислород воздуха и углеводы самого растения. Испарение воды листьями обеспечивает непрерывный ток ее с растворенными питательными веществами от корней ко всем органам надземной части куста. Установлено, что поверхность листьев в 1 м2 испаряет ежедневно до 1,5 л воды. Интенсивность испарения зависит от количества устьиц, их величины и степени открытия, а также от притока воды, температуры и влажности почвы и воздуха, наличия ветра и других факторов. Испарение снижает температуру листьев. На протяжении периода вегетации баланс фотосинтеза и дыхания у листьев по длине побега неодинаков. Верхушка побега я молодые растущие листья расходуют питательные вещества более интенсивно, чем их ассимилируют, тогда как у листьев средней и нижней зон побега ассимиляция преобладает над дыханием.
Рис. 22. Главные направления передвижения ассимилятов в побегах винограда в зависимости от фазы вегетации растения: / — в очень молодых побегах; 2 — во время цветения; 3 — после завязывания ягод (по Hale, Weaver). В начале вегетации в листьях содержится до 82% воды и накапливается небольшое количество сухих веществ (главным образом зольных). К концу периода вегетации с понижением температуры воздуха и почвы и уменьшением водообеспеченности растений процессы транспирации и фотосинтеза замедляются. Листья к этому времени начинают стареть и продуктивность их снижается. Содержание воды в них уменьшается до 70%, а общее количество золы увеличивается до 4,5—12%, причем ее больше в листьях нижнего яруса, чем среднего и верхнего. При старении листьев и прекращении из них оттока ассимилятов листья становятся тяжелыми и плотными, содержание хлоропластов, а следовательно и хлорофилла, в них уменьшается. Наибольшее количество хлорофилла содержится в листьях средней зоны побега. Осенью листья приобретают характерную окраску: у сортов с белыми и розовыми ягодами — желтую, с темными — пурпурно-красную. Окраска листьев изменяется в результате разрушения хлоропластов и красящих пигментов, а в некоторых случаях вследствие обогащения клеток антоцианом и ксантофиллом. К этому времени из листа в стебель почти полностью происходит отток органических веществ (крахмала, сахаров и др.). В листьях резко падает содержание азота и фосфора. В месте прикрепления черешка листа к побегу образуется пробковый отделительный слой, и черешок листа вместе с листовой пластинкой опадает, оставляя на побеге темный рубец. Анатомическое строение листа. С верхней стороны листовая пластинка покрыта эпидермисом, который защищен слоем плотной кутикулы, предохраняющей лист от излишнего испарения воды. Под верхним эпидермисом находится слой удлиненных клеток палисадной (столбчатой) паренхимы, содержащей много хлоропластов, которые обеспечивают фотосинтез. Под палисадной паренхимой расположена губчатая паренхима, состоящая из рыхлосоединенных клеток неправильной формы. Между ними находятся заполненные воздухом пространства — межклетники. Губчатая паренхима составляет основную толщу листа, она также содержит хлоропласта, но в меньшем количестве, чем палисадная паренхима. К губчатой паренхиме примыкает эпидермис нижней поверхности листа, где находятся устьица и выросты в виде волосков и щетинок, создающих опушение листа, и железок, выполняющих защитную роль. Почки.Образуются в пазухе листа у основания его черешка на узлах растущих побегов. При развитии почек в каждом периоде вегетации возобновляется рост побегов, поэтому почки являются точками роста. У виноградного растения различают 3 типа почек: пазушные скороспелые пасынковые, зимующие и спящие. Все они представляют собой зародышевый побег, на котором в зачаточном состоянии формируются вегетативные и генеративные органы будущего растения. Почки образуются путем деления поверхностных слоев клеток меристемы верхушки конуса нарастания. С ростом побега в пазухах листьев сначала закладываются скороспелые пазушные пасынковые почки (рис. 23). Они не имеют периода покоя, быстро формируются, и из них в тот же период вегетации развиваются побеги — пасынки. С ростом основного побега в пазухах самых нижних листьев пасынков, независимо от степени их развития (слабого или сильного), закладываются сложные зимующие почки, которые в практике виноградарства называют глазками. Из них весной следующего года развиваются основные побеги. Глазок состоит из нескольких почек — главной, или центральной, и нескольких (3–6) замещающих — запасных, меньшего размера, расположенных вокруг главной почки. Зимующая почка в течение всего периода вегетации растет, постепенно увеличивается в объеме и дифференцируется, превращаясь в глазок. В пазухах листьев пасынков закладываются также скороспелые пазушные почки. В отличие от пазушных почек зимующие глазки имеют период глубокого покоя и трогаются в рост только после его прохождения при наступлении благоприятных условий, обычно весной будущего года. Сначала трогается в рост центральная почка глазка, затем замещающие. При благоприятных условиях, а также в случае повреждения главной почки низкими температурами или другими неблагоприятными внешними факторами в рост трогается 1 либо одновременно 2–3 наиболее развитые замещающие почки. Однако развившиеся из них побеги (двойники и тройники) слабее основного и менее урожайны. Если по каким-либо причинам почки глазка (главная или замещающие) не трогаются в рост, они остаются на побеге и к осени, частично регенерируя вследствие образования пробкового камбия, постепенно углубляются в ткани многолетних частей куста. Наружная часть глазков высыхает и остается Только его внутренняя часть — почечный след, который растет вместе с ростом стебля в толщину. Такие почки условно называют спящими. Они живут долго и при благоприятных условиях — хорошем питании, малой нагрузке кустов плодовыми побегами или при гибели точек роста надземной части куста — могут пробудиться и дать побеги (порослевые и волчки), сходные по своему морфологическому строению с побегами, развившимися из пасынковой или из зимующей почки. В год развития такие побеги остаются в основном бесплодными, хотя быстро растут и утолщаются. В пазухе их листьев могут заложиться плодоносные почки, которые дадут урожай в следующем году.
Рис. 23. Развитие пасынковой почки и формирование глазка: 1 — прорастающая пазушная почка, образующая пасынок на побеге; 2 — конус нарастания пасынка; 3 — усик; 4 — прилистник; 5 — лист; 6 — первый лист пасынка; 7 — формирующаяся сложная зимующая почка (по Гордеевой). Пробуждение спящих почек и высокая побеговосстановительная способность винограда — характерные показатели установления нарушенного равновесия между развитием корневой системы и силой роста надземной части куста, его нагрузкой глазками и побегами, что позволяет в случае необходимости заново сформировать новые рукава или заменить всю надземную часть куста. На этой биологической особенности винограда основано омоложение растений (срез на черную голову) и восстановление кустов, поврежденных морозами и другими неблагоприятными факторами внешней среды. Все типы почек: пазушные скороспелые пасынковые (летние), зимующие (главная и замещающие) и спящие — по морфологическому строению одинаковы. Они различаются только степенью развития, количеством образовавшихся в них зачаточных вегетативных и репродуктивных органов будущего побега. На продольном срезе хорошо сформировавшейся зимующей почки (глазка) под микроскопом в центре можно видеть наиболее развитую главную почку, по бокам которой находятся 1–2 и больше менее развитые замещающие (рис. 24). В центре главной почки видны расширенная к основанию конусовидная ось зачаточного побега и верхушка конуса нарастания. На оси заметны поперечные чередующиеся светлые и темные полосы. Это будущие узлы и междоузлия. По обеим сторонам зачаточного стебля в виде лентовидных образований располагаются зачатки листьев, бугристые, гроздевидные зачатки соцветий, вилочковидные зачатки будущих усиков. В центральной почке, в зависимости от степени ее развития, бывает до 13 зачаточных листочков. В пазухах зачаточных листочков можно видеть бугорки зачаточных пасынковых почек. Замещающие почки имеют такое же строение, как и главная, но, поскольку они образуются и формируются позже центральной почки, отличаются от нее меньшим размером и меньшей степенью дифференциации зачаточных органов. Если на зачаточном побеге центральной почки можно насчитать до семи — восьми узлов, то замещающих — не более трех — пяти. Снаружи центральная и замещающие почки глазка покрыты Двумя крупными чешуями. Летом они зеленые, а к осени пропитываются особым веществом — субирином и пробковеют, становятся кожистыми, приобретают коричнево-бурую окраску. Под чешуями глазка находится защитный волосяной покров, предохраняющий нежные почки глазка от повреждения низкими температурами. Глазки на узлах побега расположены на небольшом плоском возвышении — подушечке. Между подушечкой и основанием глазка имеется тонкий (не более 2 мм) подстилающий слой, состоящий из тонкостенных паренхимных клеток. Ранее считали, что этот слой несет в себе зачатки почек. Однако результаты исследований этого не подтвердили. У почек винограда всех типов могут осуществляться вегетативные и генеративные процессы. Почки с зачатками соцветий называют плодоносными, листьев и усиков — бесплодным и.
Рис. 24. Продольный разрез зимующей почки (глазка) винограда: 1 — главная почка; 2 — замещающая почка первого порядка; 3 — замещающая почка. второго порядка; 4 — подстилающий слой; 5—: зачаточный лист; 6 — соцветие;, 7 — усик; 8 — чешуевидные прилистники; 9 — волосяной покров; 10 — покровные наружные чешуи; // — узел эмбрионального побега; 12 — междоузлия; 13 — внутренний паренхимный след почки (по Гордеевой). По внешнему виду отличить плодоносные почки от бесплодных у виноградного растения невозможно. Продуктивность куста зависит от количества заложившихся на побегах плодоносных почек и числа соцветий в них. Лучший период для закладки, роста и формирования почек в глазках, а внутри них — эмбриональных соцветий у большинства сортов винограда — первая половина и середина лета, в период цветения растений и после него, при условии хорошей освещенности и необходимого питания. Наиболее развитые (плодоносные) почки формируются в глазках средней части побега, в пределах 4—12–го узлов от его основания. Менее развиты почки глазка первых трех и самых верхних узлов побега. Они всегда бесплодны. В жизни эмбрионального побега (почки), как отмечал А. М Негруль (1959), имеется несколько критических периодов, в течение которых происходят существенные изменения, в частности образование и дифференциация соцветий, что и определяет, быть почке плодоносной или остаться бесплодной. Чем больше разовьется плодоносных почек, тем выше урожайность виноградных насаждений. Первый критический период при закладке и формировании почек наступает в то время, когда смещенная точка роста продолжает рост и в зависимости от условий в ней формируется соцветие или усик. У большинства сортов винограда группы бассейна Черного моря и западноевропейской зачатки соцветий в глазках основных побегов закладываются почти одновременно, обычно в конце мая — начале июня, что совпадает с периодом цветения, у сортов восточной группы — несколько позже, в конце июля. У сортов с коротким периодом вегетации закладка зачатков первых соцветии заканчивается в наиболее сжатые сроки (примерно в течение месяца), тогда как у сортов с длительной вегетацией этот период более продолжительный. У ранних сортов винограда соцветия в почках начинают закладываться на 10–14 дней раньше, чем у сортов поздних сроков созревания. Закладка соцветий в глазках пасынкового побега начинается в первой декаде июля сначала на пасынках, развившихся в нижней и средней зонах основного побега, с 1–4–и 7–9–го узлов, а в конце июля — на пасынках, развившихся в верхней зоне основного побега. Степень плодоносности почек по длине побега неодинакова. Наиболее плодоносны и лучше дифференцированы соцветия в глазках, расположенных в пределах 5—10–го узлов. От закладки бугорка до дифференциации первого соцветия проходит примерно 22–23 дня, второго соцветия—1$—30 дней, закладка и дифференциация которого начинается на 10–12 дней позже, чем первого соцветия. Максимальное количество соцветий закладывается в фазе цветения, которая считается вторым наиболее критическим периодом в годичном цикле развития виноградного растения. В фазе роста ягод закладка и дифференциация соцветий продолжаются, но в меньшей степени. Самые крупные соцветия закладываются в более развитых почках глазков средней зоны побега, в пределах 5—10–го узлов от его основания, что объясняется наиболее благоприятными внешними условиями — достаточно высокой температурой и влажностью воздуха, хорошей освещенностью, активной ассимиляцией листьев как основных, так и пасынковых побегов. Все это обеспечивает хорошее питание почек, а следовательно, их закладку и дифференциацию. Зачатки соцветий у виноградного растения могут закладываться в центральной и замещающих почках глазка, в скороспелых пасынковых почках, а иногда и в спящих почках при условии: оптимального водно-воздушного режима, обеспеченности питательными элементами и хорошей освещенности кустов, что достигается выбором хорошо прогреваемых открытых участков, конструкций насаждений, внесением удобрений, орошением виноградников и. Другими приемами по уходу за растениями и почвой. Соцветие, усик, цветок. Зачаточные соцветия имеют вид полукруглых бугорков на узлах эмбрионального побега. В росте соцветий наблюдается определенная ритмичность. В самом начале вегетации их рост коррелирует с ростом побегов, и чем сильнее Рост зеленых побегов, тем сильнее рост соцветий. Затем он несколько затухает, а ко времени цветения полностью приостанавливается. К этому времени соцветия достигают 50–60% своей Длины. В период массового цветения наблюдается новая волна роста соцветий, максимум которого приходится на период, когда ягоды достигнут величины горошины. Временная остановка роста соцветия в период цветения символизирует собой как бы переход из одного качественного состояния органа в другое — соцветия в гроздь. С ростом и развитием соцветий в течение всего периода вегетации идет процесс их дифференциации, который не заканчивается осенью, а продолжается весной следующего года. Соцветие растет от основания к вершине. Полностью развитое соцветие винограда — сложная кисть или метелка, имеющая конусовидную форму. В 1–ом различают ножку, с помощью которой оно прикрепляется к побегу, и центральную ось с разветвлениями I-, 2-, 3-, реже 4–го порядка. На их концах находятся бутоны, собранные в группы по 3 шт. Центральный бутон развит всегда сильнее, чем 2 боковых. Пока соцветие растет, все его части остаются зелеными. Развиваются соцветия на вегетирующем побеге в пределах 4—12–го узлов с противоположной листу стороне. На одном побеге в зависимости от сортовых особенностей и факторов внешней среды может быть от 1 до 5–7 соцветий. Соцветия различаются как по величине, так и степени развития. У сортов с мелкими гроздями в соцветиях насчитывается 50—200, с крупными — 700—1500 бутонов. До начала цветения соцветие растет очень интенсивно, достигая 50–60, к концу цветения — 75% своей нормальной величины. Между ростом побега и соцветия существует корреляция: чем сильнее рост побега, тем более интенсивен рост соцветий. Соцветия, расположенные ближе к основанию плодоносного побега, всегда крупнее вышерасположенных. Соцветия могут быть с усиком и усиковые, что определяется внутренними и внешними факторами, в которых они закладывались и формировались (рис. 25). От величины соцветия и количества завязавшихся в нем ягод в значительной степени зависит размер грозди и величина будущего урожая. Нередко на побегах винограда можно видеть переходные формы от соцветия к усику и усика к соцветию (см. рис. 25). Изучение морфологии этих органов подтверждает их единое стеблевое происхождение. Встречаются аномалии в строении соцветий, выражающиеся в фасциациях, образовании широких и плоских осей соцветий и пучков бутонов с укороченными цветоножками. Усики имеют стеблевое происхождение и представляют собой метаморфизованное соцветие. Располагаются усики на узлах побега против листьев. На плодоносных растущих побегах усики находятся на узлах выше соцветий, на бесплодных — по всей длине побега, начиная со 2–5–го узла. При помощи усиков побеги прочно прикрепляются к опорам — деревьям, кольям, проволокам шпалеры и др. В поисках опоры верхушка усика, как установил еще Ч. Дарвин, совершает круговое (нутационное) движение, делая в течение 2 ч полный круг, и при соприкосновении с предметом под влиянием раздражения закручивается вокруг опоры, а свободная нижняя часть усика приобретает вид пружины и таким образом подтягивает побег к опоре. У прицепившихся к опоре усиков развивается механическая ткань (либриформ), они древеснеют и становятся очень прочными. Если же при круговом движении усик не встречает опоры, то он некоторое время остается травянистым, зеленым, а затем вскоре засыхает и отпадает. Усики бывают простые и разветвленные — двойные, сильно разветвленные с бутонами. Усик ветвится так же, как и побег. Нередко в месте разветвления на усике образуются листья, иногда сам усик превращается в побег с листьями и соцветиями. Такие побеги называются внепазушными. Анатомическое строение осей соцветия и усика аналогично первичному анатомическому строению молодого побега. Цветок винограда, как у всех семенных растений, относится к репродуктивным органам и выполняет функцию семенного размножения. У виноградного растения цветки невзрачные, мелкие, желто-зеленые, сидят на тонких ножках. Рис. 25. Переходные формы усика к соцветию: а — внепазушный побег; б — переходы от внепазушного побега к усику; в — от усика к соцветию.
Чашечка недоразвитая, состоит из 5 сросшихся недоразвитых чашелистиков, охватывает основание цветка слабовыраженными тупыми зубчиками. Венчик состоит из 5 сросшихся вверху лепестков, прикрывающих внутренние части цветка. Между чашечкой и основанием венчика находится небольшой кольцевой валик из недоразвитых нектарников, окрашенных в зависимости от сорта в зеленый, желтый либо в оранжевый цвет. В них содержатся эфирные масла с приятным запахом, напоминающим запах резеды, но. нектара они не выделяют, поэтому и не привлекают пчел. Тычинок в цветке 5, реже 6–7. Они расположены против лепестков венчика и слегка изогнуты. На конце тычиночных нитей находятся пыльники, содержащие пыльцу. Пыльники 2–гнездные, с широким связником, прикрепленным основанием к тычиночной нити. В каждом гнезде по 2 пыльцевых мешка, которые во время цветения открываются продольными щелями, в результате чего из них высыпается пыльца. Пестик бутылкообразной или грушевидной формы, состоит из завязи, столбика и рыльца. Завязь верхняя, 2–гнездная, состоит из двух плодолистиков. В одном гнезде 2 анатропные семяпочки. В каждой из них находится многоклеточное ядро-нуцеллус с двумя покровами (интугементами) — внутренним и внешним, между которыми находится узкий канал — пыльцевход. В нуцеллусе семяпочки из археспоральнои клетки развивается зародышевый мешок, содержащий яйцеклетку, синергиды, полярные ядра и антиподы. Наряду с общей схемой строения (2–гнездная завязь с двумя семяпочками в каждом гнезде) у многих сортов встречаются отклонения (1–8%), выражающиеся в наличии большого числа гнезд в завязи и семяпочек в них. Этим объясняется наличие более четырех семян в ягоде. Столбик пестика цветка у разных видов и сортов винограда может быть коротким и толстым, удлиненным и тонким. Рыльце блюдцеобразной формы, с беловатыми приподнятыми ровными, реже вальчатыми краями. У винограда несколько типов цветков (рис. 26): обоеполые (облигатно-фертильные); функционально-женские (факультативно-фертильные); истинно женские и мужские (облигатно-бесплодные). У дикого винограда 2 основных типа цветка: мужской с различными вариациями и женский. Большинство культивируемых сортов европейско-азиатского винограда имеют обоеполый тип цветка. Как исключение в старых насаждениях встречаются сорта с функционально-женским, в насаждениях маточников филлоксероустойчивых подвоев — сорта в основном с мужским типом цветка. Все названные типы цветка различаются по морфологическим признакам. От умения распознавать тип цветка зависит правильное размещение сортов в насаждениях и организация работ по искусственному опылению самостерильных (бесплодных) сортов, урожай и качество винограда.
Рис. 26. Тип цветка винограда: а - обоеполый; б — функционально-женскнй; в — истинно женский; г — мужской. Цветок обоеполого типа имеет прямостоячие тычинки, которые выше пестика или равны ему. В пыльниках находится жизнеспособная (фертильная) пыльца. Завязь хорошо развита, обычно грушевидной формы. Пыльца обоеполого цветка в сухом виде имеет правильную удлиненно-бочкообразную форму, в случае погружения в 10–15%-ный раствор сахара (глюкозы) при температуре 25–30°C легко прорастает в пыльцевую трубку. Сорта винограда с обоеполым типом цветка в основном самоопыляющиеся (автогамия), но встречаются сорта, нуждающиеся в перекрестном (аллогамия) опылении. Цветок функционально-женского типа также имеет пестик и тычинки, но по морфологическому строению отличается от обоеполого. Тычинки менее развиты, короче пестика, пестик довольно развитый. Во время цветения после сбрасывания колпачка тычинки отгибаются книзу. Пыльца бесплодная — стерильная, в сухом виде имеет неправильную форму: угловатую, ромбическую, трехгранную, округлую, овальную, шаровидную и т. д. В растворе сахара пыльца цветка этого типа не прорастает в пыльцевую трубку, а только едва набухает и принимает округлую форму. Сорта с таким типом цветка самостерильны и нуждаются в перекрестном опылении. Бесплодность пыльцы и недостаточное искусственное опыление у сортов с функционально-женским типом цветка — причины чрезмерного осыпания цветков, а иногда и засыхания соцветий. Если сорта винограда с функционально-женским типом цветка своевременно не опылить, то ягоды на них или не завязываются, или бывают мелкими, величиной с горошину (партенокарпические), без семян либо с мелкими Щуплыми семенами без зародыша и эндосперма. Мужские цветки винограда имеют сильноразвитые прямостоячие тычинки с большим количеством жизнеспособной пыльцы в пыльниках, завязь недоразвита, рыльце на пестике отсутствует, сам пестик имеет различную форму, от маленького бугорка до сильноразвитой крупной завязи. Пыльца цветка мужского типа, как и у обоеполого, в сухом виде имеет правильную удлиненно-бочкообразную форму. Она также хорошо прорастает в растворе сахара в течение нескольких часов в пыльцевую трубку. Растения с мужским типом цветка имеют крупные соцветия, но ягод не образуют. В том случае, если по внешним морфологическим признакам трудно определить тип цветка, его устанавливают по форме и способности прорастания сухой пыльцы. Соцветия с мужским типом цветка используют для сбора пыльцы, необходимой для опыления сортов винограда с функционально-женским типом цветка. Есть и очень ценные старые сорта, такие как Нимранг, Чауш белый, Пухляковский, Тавквери, Катта-Курган и другие, с функционально-женским типом цветка, нуждающиеся в перекрестном опылении. В посадках винограда к таким сортам подбирают соответствующие опылители, у которых совпадает срок цветения. Их высаживают рядами — 1 ряд сорта — опылителя и 2— опыляемых. Кроме того, применяют искусственное опыление. Однако из-за технологических сложностей возделывания и снижения урожайности в годы с неблагоприятными для опыления погодными условиями новые сорта с таким типом цветка Государственной комиссией по сортоиспытанию сельскохозяйственных культур Госагропрома СССР в настоящее время не принимаются. Помимо основных указанных типов цветка, у винограда встречаются и истинно женские цветки (клон сорта Мурведр), а также различные аномалии в строении цветка. Чаще всего аномалии проявляются в раскрывании цветка розеткой, звездочкой или в махровости. Махровость присуща в основном сортам, имеющим функционально-женский тип цветка. Встречаются и фасциации цветков, выражающиеся в слиянии двух цветков в один сложный или в появлении цветков со сросшимися двумя — тремя цветоножками в одну и др. Гроздь, ягода, семя.Соцветие после оплодотворения и естественного опадения цветков и завязей растет и формируется в гроздь. Ножка соцветия преобразуется в ножку грозди, ось соцветия с разветвлениями — в гребень, завязи — в ягоды. В процессе роста и развития гребня в нем наиболее сильно развиваются механические и проводящие ткани, в частности мягкий луб, что обеспечивает прочность гребня и приток к ягодам большого количества пластических веществ. Ножка грозди, при помощи которой гроздь прикрепляется к побегу, в зависимости от сорта бывает короткой и длинной. У одних сортов ножка все время остается травянистой, зеленой, а у других она древеснеет. На ножке, недалеко от места ее прикрепления к побегу, имеется утолщение — узел, от которого ответвляется усик. Если усик имел на себе цветки, то после их опыления усик превращается в ответвление основной грозди, называемой крылом. У некоторых сортов их может быть 1–2. Крылья отличаются от основной грозди меньшей величиной и меньшим количеством ягод. Иногда крылья развиваются наравне с основной осью грозди. Разветвленная часть гребня заканчивается плодоножками, на которых сидят ягоды. От характера ветвления гребней и длины осей 1-, 2–го и последующих порядков зависят величина, форма и плотность грозди, которые являются сортовыми признаками и имеют важное практическое значение.
Рис 27. Строение ягоды винограда, продольный разрез: 1. плодоножка; 2 — подушечка; 3 — сосудистые пучки; 4 — семена 5 —кожица; 6–мякоть; 7 — сердечко; 8 — эпидермис. Эти показатели могут изменяться в зависимости от почвенно-климатических условий и приемов агротехники. Однако грозди, расположенные ближе к основанию плодоносного побега, всегда крупнее вышерасположенных. На рост гроздей и ягод винограда значительно влияет обработка гиббереллином и другими регуляторами роста. Величина грозди определяет урожайность сорта и удобство ручной уборки. Чем больше средняя масса грозди, тем выше урожай с 1 га насаждений. Анатомическое строение ножки грозди и плодоножки ягоды аналогично строению междоузлия молодого побега. Ягода — основной орган, ради которого культивируют виноград. Она развивается из завязи цветка. После оплодотворения яйцеклетки образуется зародыш, который стимулирует рост завязи (околоплодника). Рыльце и пестик высыхают, превращаясь в пупок, хорошо заметный на вершине вызревшей ягоды. К гребню грозди ягода прикреплена с помощью плодоножки, расширенной у основания в подушечку (рис. 27). Через плодоножку и подушечку из гребня в ягоду проходят сосудисто-волокнистые пучки, которые в виде мелкой сетки расходятся под кожицей мякоти ягоды и идут к семенам. При отрыве ягод от плодоножки на подушечке остается кисточка. Это оторванные сосудисто-волокнистые пучки. Кожица ягод (эпикарпий) состоит из однослойного эпидермиса толщиной 30–40 мкм, клетки которого напоминают правильный многоугольник, и 10–15 слоев клеток кожицы, расположенных под эпидермисом. В эпидермисе имеются щели — устьица, через которые происходит газообмен. Сверху клетки эпидермиса утолщены и кутикулонизированы. Количество слоев кожицы, их толщина и плотность обусловливают устойчивость ягод к загниванию, а у столовых сортов — транспортабельность и пригодность для длительного хранения. С ростом ягоды клетки кожицы растягиваются, становятся округлыми и переходят в радиально вытянутые клетки мякоти — мезокарпий. Это основная часть ягоды клетки мякоти не прилегают друг к другу. Между ними имеются большие сообщающиеся между собой межклеточные пространства, через которые осуществляется газообмен. В мякоти выделяются 2 зоны. Первая зона находится непосредственно под кожицей и состоит из нескольких слоев крупных радиально распложенных клеток, достигающих 0,3–0,4 мм в диаметре. Вторая зона — это самый внутренний, прилегающий к семенам слой (эндокарпий, сердечко), состоящий из 5–6 слоев удлиненных клеток. Общее число слоев клеток ягоды от эпидермиса до эндокарпия такое же, как у завязи (около 25–30). У столовых сортов винограда количество слоев клеток кожицы и сами клетки больше, ко времени созревания они сильно растягиваются. Клетки мезокарпия, прилегающего к кожице, имеют плотную мембрану и выделяют небольшое количество сока, что обусловливает их характерную мясистую, мясисто-сочную, плотную хрящеватую, хрустящую консистенцию. У столовых сортов кожица не отделяется от мякоти и при нагрузке раздавливается вместе с ней. У технических сортов число слоев клеток кожицы меньше и сами клетки мельче. Мезокарпий же состоит из более крупных клеток с очень нежными мембранами, которые при созревании винограда растворяются, вследствие чего мякоть ягод представляет сплошную жидкую массу. Кожица у большинства винных сортов тонкая, эластичная, легко разрывающаяся, с большим количеством сока в вакуолях, легко отделяется от мякоти при раздавливании. Оболочка клеток мякоти, представленная клетчаткой и целлюлозой, составляет 0,3–0,5% массы мякоти, остальное — это клеточный сок, содержащий у зрелых ягод до 55–95% воды, 5—30— сахаров (глюкоза, фруктоза, сахароза), 0,5–1,9% органических кислот (винная, яблочная, глюконовая, глюкуроновая, лимонная и др.). Витамины (А, Вь В2, РР, С) содержатся в основном в кожице. В мякоти и соке их меньше. Дубильные вещества, так же как крахмал, имеются в мякоти только до начала созревания ягод, затем их количество резко уменьшается и ко времени созревания ягод они полностью исчезают. Кроме того, в ягодах накапливаются ароматические вещества, которые представляют собой сложный комплекс летучих ароматических эфирных масел, обусловливающих аромат ягод. Они накапливаются вблизи проводящих пучков, примыкающих к кожице. Пока ягода растет, она дышит, ассимилирует и остается зеленой. В ней содержатся хлорофилл и красящие вещества — каротин, ксантофилл и др. Красящие вещества накапливаются в ягоде постепенно. Максимальное их количество наблюдается в период полной зрелости ягод. В процессе созревания ягод вещества, обусловливающие их первоначальную окраску, разрушаются, и в клетках кожицы, в одном, двух или во всех ее слоях, накапливаются другие пигменты — флавонолы, основные компоненты которых— кверцитин и кверцинтрин, обладающие желтой окраской. Это продукты изменения хлорофилла, каротина, ксантофилла и других сопутствующих хлорофиллу веществ. Они придают сортам с белой ягодой окраску от светло-зеленой до золотисто-янтарной. Другие пигменты, представляющие антоциановый комплекс, в состав которого входят ампелопсин, энин и прочие, обусловливают образование красных веществ, придающих ягодам розовую, красную и черно-синюю окраску. Цвет ягод — четкий ампелографический признак. Однако в зависимости от экологических условий выращивания винограда у одного и того же сорта он может изменяться: быть светлее или темнее. Так, например, у сортов Тайфи розовый, Октябрьский, Забалканский в условиях Крыма ягоды приобретают интенсивно-розовую и даже красную окраску, тогда как в Средней Азии ягоды у этих сортов светло-розовые. В клетках мякоти красящих веществ обычно не бывает, но есть сорта (Пти Буше, ВИР 1, Одесский черный, Саперави и др.), красящие пигменты у которых находятся и в вакуолях клеток мякоти. Это так называемые сорта — красильщики, из которых можно приготовить только интенсивно окрашенные соки и вина. На поверхности кожицы, особенно у сортов со светлой окраской ягод, заметны коричневые точки — чечевички, или остатки опробковевших ко времени созревания ягод устьиц. Снаружи кожица ягод покрыта восковидным сизо-голубоватым налетом — пруином, наиболее ярко выраженным у темноокрашенных сортов. Пруин выполняет защитную роль: предохраняет ягоду от неблагоприятных внешних условий, потери воды, гниения. Ко времени созревания кожица ягод в зависимости от сорта становится тонкой, легко рвущейся, прозрачной, не ощущаемой при еде или толстой, грубой и очень плотной. Ягоды разных сортов винограда различаются по величине, форме, окраске и вкусовым качествам. Мякоть ягоды может быть сочной, тающей, плотной, хрустящей, мясистой, слизистой. У сортов винограда с сочной мякотью ягод стенки клеток тонкие, легко разрываются при раздавливании, и сок при этом быстро вытекает. Сорта с такой ягодой нетранспортабельны и не хранятся. Наоборот, у сортов с плотной хрящевой мякотью ягоды менее сочные и при раздавливании плохо отдают сок. Такие сорта пригодны для длительного хранения и транспортировки на дальние расстояния. Вкус ягод определяется гармоничным сочетанием в мякоти сахара и кислоты. Ягоды многих сортов винограда имеют специфический аромат (группа мускатных сортов — Мускат белый, Мускат черный, Мускат венгерский, а также сорта Рислинг, Траминер розовый, Каберне Совиньон, Изабелла, Лидия). Некоторые сорта (группы Шасла, Португизер и др.) выраженного аромата не имеют. Величина (масса) ягоды зависит от числа семян в ней. С увеличением числа семян от одного до четырех увеличивается масса ягоды. На каждое семя приходится около 10% массы ягоды. От того, как прошло оплодотворение и сколько развилось семян в ягоде, зависит масса ягоды и всей грозди, а следовательно, и величина урожая с куста и единицы площади. Ягоды у винограда бывают с семенами и без семян. Бессемянных сортов немного. Из общего числа районированных в СССР сортов винограда (свыше 250) бессемянных только 9, или около 4%. Согласно схеме происхождения бессемянных сортов винограда, разработанной и предложенной К. В. Смирновым, первые бессемянные формы винограда появились в виде мутаций на семенных сортах. Затем в результате естественной гибридизации между семенными и бессемянными сортами возникли вторичные бессемянные формы. Путем целенаправленной селекции сортимент их был расширен. Бессемянные сорта делят на 2 группы: коринок (белая, розовая, черная), у которых образование и рост ягод проходят за счет разрастания завязи до величины горошины, без процессов опыления и оплодотворения (партенокарпия); кишмишей (белый, розовый, черный и новые бессемянные сорта), которым для образования ягод необходим процесс опыления. Процесс же оплодотворения нарушается на различных этапах развития, что приводит к образованию мелких недоразвитых семян (рудиментов), значительно различающихся по величине и степени развития (стеноспермокарпия). У отдельных бессемянных сортов и гибридных форм может встретиться 2 формы бессемянности — партенокарпия и стеноспермокарпия. Так, у клонов Кишмиша белого в одной грозди можно одновременно обнаружить ягоды бессемянные, овальной формы, свойственной сорту величины (кишмишность) и круглые, мелкие (партенокарпия). Имеет место и частичная партенокарпия, когда в нормально развитой грозди с развитыми семенами в ягодах часть ягод остаются бессемянными, недоразвитыми (горошащимися), что наблюдается при плохом опылении сортов с функционально-женским типом цветка (Мадлен Анжевин, Чауш белый и др.). В этом случае в процессе оплодотворения пыльцевые трубки достигают нуцеллуса, но не входят в него, а переплетаются между собой, образуя плотный клубок, и в таком виде отмирают. Семена у винограда мелкие, грушевидной формы, с заостренным удлиненным клювиком. Пока ягода растет, семена находятся в молочной зрелости, мягкие по консистенции и зеленые по цвету. Ко времени созревания семена покрываются прочной кожурой, содержащей каменистые клетки, богатые фенольными веществами. Благодаря этому они становятся твердыми и приобретают желтовато-коричневую окраску. Под кожурой семени находится эндосперм, клетки которого богаты питательными веществами — белком, жиром. В клювике семени расположен зародыш, несущий зачатки органов будущего растения: корешок, обращенный к микропиле, стебелек, 2 семядоли сердцевидной формы, между которыми расположена зародышевая верхушечная почечка — эпикотиль. Семя винограда имеет брюшную и спинную сторону. На брюшной стороне 2 продольные бороздки — впадины, между которыми проходит семенной шов, переходящий на более выпуклую округлую спинную сторону до халазы — места проникновения в семя сосудисто-волокнистых пучков. В ягодах содержится обычно 2–3 семени, но бывает 4 и более. Как указывалось выше, есть и бессемянные сорта винограда, которые, согласно классификации К. В. Смирнова, по величине рудиментов семян и степени их развития делят на 3 категории. К первой категории относят сорта винограда, в ягодах которых содержатся мелкие (до 6 мг) рудименты семян (Кишмиш белый круглый, Кишмиш белый овальный, Кишмиш розовый, Кишмиш черный), ко второй — сорта с рудиментами семян 6,1 —10 мг (Аскери, Бедона), к третьей категории — с более крупными рудиментами— 10,1 — 14 мг (Кишмиш люнда и др.). Семена разных видов и сортов винограда различаются по величине, форме, окраске, длине клювика, расположению и форме халазы. У сортов V. vinifera семена крупные и имеют удлиненный клювик. Халаза расположена в верхней трети спинной стороны, тогда как у других видов — в ее средней части. Кожура семени состоит из трех, слоев — наружного, промежуточного и внутреннего. Наружный слой имеет один ряд тангентально удлиненных клеток, промежуточный — несколько рядов тонкостенных клеток рыхлого строения, служащих у невызревших семян вместилищем крахмала и рафид, а у вызревших они высыхают, сплющиваются и становятся местом накопления танина. Внутренний, защитный слой примыкает к эндосперму и состоит из двух — шести рядов радиально удлиненных каменистых клеток с одревесневшими стенками. Оболочка из каменистых клеток полностью окружает семена винограда, кроме халазы и верхушки клювика, где находится отверстие — микропиле. В клювике каменистых клеток значительно больше, чем в остальной части семени. Большой жизненный цикл и возрастные этапы развития виноградного растения в онтогенезеЖизнь виноградного растения — многолетней древесной лианы — в естественных условиях длится многие десятки лет. На протяжении этого периода оно растет и развивается, подчиняясь общим биологическим законам, суть которых сводится к гармоничному сочетанию морфологических изменений, биологических особенностей и физиологических функций. Ч. Дарвин впервые обосновал и доказал наличие четкой зависимости индивидуального развития организма в его жизненном цикле, то есть онтогенеза, от филогенеза — процесса исторического эволюционного развития вида. Онтогенез растений — это совокупность закономерных морфологических и физиологических изменений, протекающих в организме от его зарождения из полового или из вегетативного зачатка До естественного отмирания и возникающих на основе исторически сложившейся природы данного организма в конкретных условиях внешней среды. Онтогенез охватывает большой и малый жизненные циклы виноградного растения. В процессе индивидуального развития происходит формирование организма и качественные изменения новых клеток, органов или их частей, которые в процессе роста увеличиваются в размере, что приводит к нарастанию массы растения. В зависимости от способа размножения винограда — семенного или вегетативного (отрезками стебля) — развитие растений в онтогенезе протекает неодинаково. У растений, выращенных из семян (сеянцев) в их индивидуальном развитии (онтогенезе), большое значение имеет наследственность — свойство организма передавать при размножении потомству признаки, приобретенные родителями, и изменчивость — способность организма приобретать новые признаки или утрачивать старые как под влиянием условий внешней среды, так и при воздействии на растения различными факторами среды, мутагенами и др. У растений, размноженных вегетативным путем, в потомстве сохраняются все наследственные признаки материнского растения и они не подвергаются изменчивости, свойственной сеянцам винограда. Растение винограда в своем большом цикле развития от прорастания семени у сеянца и распускания глазков у вегетативно размноженных растений до отмирания во взрослом состоянии проходит возрастные этапы, при которых наследственные факторы действуют особенно активно и определяют признаки организма. В общебиологическом понимании этапность онтогенеза — морфологическая и функциональная расчлененность, проявляющаяся в изменении характера роста, дифференциации и функциональной активности организма. Это свойство обозначают также как дискретность, фазность или расчленимость онтогенеза. М. X. Чайлахян выделяет 5 этапов в большом жизненном цикле растений: эмбриональный, ювенильный, зрелость, размножение и старость. У многолетних плодовых культур и винограда принято выделять 4 возрастных этапа: эмбриональный, ювенильныи, продуктивный и этап отмирания. Первый этап-эмбриональный.У растений, выращенных из семян, он начинается со слияния мужской и женской половых клеток, образования зиготы, формирования зародыша и созревания семени. При благоприятных условиях тепла и влаги в семени начинаются активизироваться жизнедеятельные процессы — дыхание, превращение с помощью ферментов нерастворимых соединений в растворимые, поступление питательных веществ из клеток эндосперма в зародыш, что способствует прорастанию семени, появлению у него семядольных листьев и одного настоящего листа. Этим завершается эмбриональный этап сеянца. Продолжительность его от 6 месяцев до 2,5 года. В это время растение наиболее пластично и восприимчиво к различным факторам внешней среды. У вегетативно размноженных растений эмбриональный этап длится от начала закладки бугорка почки в пазухе листьев до ее прорастания. Этот этап протекает значительно быстрее, чем у сеянцев. Второй этап-ювенильный (юношеский).Начинается с образования у сеянцев первых настоящих листьев и интенсивного роста всех органов растения. На этом этапе наблюдается активное развитие корневой системы и накопление большого количества органических веществ во всех органах растения. В нем происходят сложные качественные превращения, без чего невозможен следующий этап развития. В ювенильный период виноградное растение очень чувствительно к условиям внешней среды, нуждается в усиленном питании и обеспеченности влагой, поэтому вся агротехника должна быть направлена на создание условий, способствующих активному росту надземных органов виноградного растения и его корневой системы. При обычном посеве семян в грунт растения начинают плодоносить на 3–5–й год, а сорта восточной группы — на 4–7–й год. У вегетативно размноженных растений этот этап значительно короче, поскольку в черенке, глазке уже прошли первые качественные этапы эмбрионального и ювенильного развития и все части потенциально готовы к плодоношению. Но оно начинается только на 2–3–й год, когда сформируются многолетние части куста (штамб, рукава). Управление этим этапом имеет важное практическое значение: у семенного потомства — приближение срока вступления в плодоношение и сокращение селекционного процесса, у вегетативного-ускоренное вступление в плодоношение. У сеянцев этот этап можно сократить путем создания обильного питания (прививка сеянца в фазе семядольных листьев на взрослое плодоносящее растение, выращивание сеянцев методом гидропоники), а также путем продления периода вегетации (использование защищенного грунта). У вегетативно размноженных растений (саженцев) приблизить срок вступления их в плодоношение можно за счет высокого уровня агротехники, в том числе повышенного фона питания молодых растений. Окончанием ювенильного этапа считают вступление растений в плодоношение. Третий этап-продуктивный, или возмужалости.В виноградарстве его называют периодом плодоношения. Это наиболее продолжительный этап, характеризующийся гармоничным сочетанием ростовых и генеративных процессов, максимальным развитием надземной и корневой систем растения, обильным плодоношением, Достигающим максимума как у сеянцев, так и у вегетативно размноженных растений к 10–20 годам. В этот этап морфологические и биологические признаки, а также характер плодоношения сеянцев приобретают постоянство. В практике возделывания винограда, выращенного вегетативным путем, стремятся максимально удлинить период плодоношения. Это достигается применением комплекса агротехнических приемов по уходу за почвой и кустом, обеспечивающих выращивание мощных, здоровых растений. К таким приемам относятся: удобрение, орошение, правильная нагрузка кустов глазками и побегами, регулируемая обрезкой, обломкой и др. При правильном соблюдении агротехники у винограда не должна появляться периодичность плодоношения. Мощные кусты более долговечны и дольше плодоносят. Период плодоношения на таких виноградниках начинается обычно со 2–3–го года и продолжается 40–50 лет. Затем их урожайность снижается, и насаждения становятся нерентабельными. В этих случаях проводят омолаживание кустов или перезакладку насаждений. В конце третьего этапа затухает рост, снижается плодоношение и наступает последний, заключительный этап жизни растения. Четвертый этап-отмирание.На этом этапе проявляются признаки старения. По современной гипотезе, организм как целое начинает стареть после того, как он исчерпает свою основную физиолого-биологическую функцию — воспроизводство нового поколения. Общебиологические основы старения состоят в разнообразных нарушениях функций белков и нуклеиновых кислот. Его следует рассматривать как систему внутренней организации организма в целом, его тканей и органов, а также молекул и органоидов клетки. Старение растений начинается с верхушек побегов и ветвей высшего порядка ветвлений, наиболее удаленных от корней. Процесс старения выражается в постепенном ухудшении взаимодействия между корневой системой и надземной частью куста. В начальном периоде онтогенеза взаимодействие между корнями и фотосинтезирующими органами наиболее полное, по мере старения оно ослабляется. Из факторов внешней среды наиболее сильно воздействует на процесс старения недостаток воды, повышенная температура воздуха и почвы, нехватка в почве азота и калия, избыточное поглощение растением кальция. Важнейшие показатели старения — снижение оводненности тканей, содержания белков, фотосинтеза и дыхания, изменение направленности действия ферментов в сторону гидролиза вместо синтеза, появление в клетках сосудисто-проводящей системы отложений — тиллов, закупоривающих сосуды. Эти изменения вызывают образование мертвой древесины внутри многолетних частей куста, сначала рукавов, а затем головы и штамба. Наиболее долговечные части куста — корни и надземный штамб. Это основные органы, используемые при омолаживании насаждений. В рукавах же процессы старения проходят быстрее, и, как правило, для восстановления жизнедеятельности кустов и хорошего роста побегов старые, поврежденные, чаще всего некрозом, рукава удаляют, заменяя их новыми из побегов, выращенных из спящих почек на штамбе или на голове куста. В рукавах сосуды функционируют 2–5 лет, иногда 6–7, в подземных штамбах — 8—10, в скелетных корнях—13–15 лет и более. Нанесение ран на многолетних частях куста ускоряет процесс старения, хотя у растения появляется тенденция к усилению процессов восстановления. Однако это увеличивает некроз тканей, приводящий к образованию мертвой древесины в подземном штамбе и старых корнях. Со временем затухают и процессы восстановления, после чего куст погибает. Продолжительность жизни каждого растения зависит от биологических особенностей сорта, силы его роста (сильнорослые сорта более долговечны), экологических условий и агротехники. В отличие от других многолетних растений виноград имеет некоторые отличительные особенности в прохождении своего большого жизненного цикла. Ему свойственна последовательность (этапность) в характере изменения морфологических признаков и выполняемых растением физиологических функций. Малый годичный цикл развития виноградаОдновременно с большим жизненным циклом виноградное растение ежегодно проходит малый годичный цикл своего развития, который охватывает 2 основных периода: покоя и вегетации. Период покоя. Не следует понимать как абсолютное отсутствие проявления жизненных процессов в растении. В клетках его органов в этот период происходят, хотя и очень медленно, сложные физиологические процессы, связанные с углеводным обменом, дыханием, испарением и химическим превращением накопленных в растении органических веществ, в частности гидролиз запасных углеводов и прежде всего крахмала, превращения его в сахара, снижение количества углеводов и др. Период относительного покоя слагается из трех фаз: условного (подготовительного) покоя; органического (физиологического) глубокого покоя и зимнего (вынужденного) покоя. Фаза условного (подготовительного) покоя начинается осенью с подготовки растения к зиме, по окончании вызревания однолетних побегов, когда они приобретают присущую сорту окраску. В этой фазе почки глазков при нормальных погодных условиях не развиваются, однако, если растение или черенки от него поместить в благоприятные условия тепла, влажности и освещенности, почки глазка могут быстро тронуться в рост. Фаза органического (физиологического) глубокого покоя определяется состоянием непрорастания почек глазка. Глубокий покой почек наступает еще до окончания периода вегетации с нижних глазков побега и длится в южной и средней зонах виноградарства в сентябре — октябре (1,5–2 месяца), когда у растений еще продолжаются процессы роста, ассимиляции, вызревания побегов и накопления в них питательных веществ (рис. 28). Фазу органического покоя следует рассматривать как приспособление растения к условиям перезимовки, препятствующее развитию почек в период осенних оттепелей, когда рост побегов прекратился, а жизнедеятельность корней продолжается. Выход почек глазка из состояния физиологического покоя У сортов различных эколого-географических групп неодинаков. Наибольшую продолжительность состояния глубокого покоя имеют почки глазка у сортов западноевропейской группы и бассейна Черного моря (Ркацители, Каберне, Каберне Совиньон), наименьшую— восточной группы (среднеазиатские сорта — Хусайне белый, кишмиши). Затем наступает фаза зимнего (вынужденного) покоя, при котором состояние непрорастания почек обусловлено неблагоприятными условиями внешней среды, в основном низкими температурами. При зимнем покое у растений отсутствуют листья и прекращается рост. Вынужденный покой может длиться 3–4 месяца и более, с ноября до марта. Но если в период вынужденного покоя с растений нарезать черенки и поместить в воду в теплом помещении, почки на побегах выйдут из периода вынужденного покоя и начнут распускаться. Растения могут быть выведены из вынужденного покоя и зимними оттепелями. Корни виноградного растения не имеют периода покоя. Они и в зимний период продолжают поглощать из почвы воду с растворенными минеральными солями, пополняя их запасы в растении, в котором в течение всей зимы проходят многие физиологические процессы — дыхание, транспирация и др. Период вегетации.В районах виноградарства с субтропическим и умеренно теплым климатом в период вегетации растение наиболее активно проявляет свои жизненные функции: наблюдается интенсивная деятельность корней, распускание почек, рост и развитие вегетативных и генеративных органов, фотосинтез, способствующий накоплению органических веществ зелеными частями растения, плодоношение, а также процессы дыхания, транспираций.
Рис. 28. Динамика покоя почек глазка винограда (по Кондо). Период вегетации у виноградного растения условно делят на (6 следующих фаз: Первая фаза — сокодвижение («плач» винограда), от начала весеннего сокодвижения до начала распускания почек глазков; вторая фаза — рост побегов и соцветий, от начала распускания почек до цветения; третья фаза — цветение, от начала до конца цветения и завязывания ягод; четвертая фаза — рост ягод, от начала образования завязей до начала созревания ягод; пятая фаза — созревание ягод, от начала созревания ягод до полной (физиологической) зрелости; шестая фаза — листопад, от полной зрелости ягод до опадения листьев и вызревания побегов. Каждая из этих фаз характеризуется определенными морфологическими изменениями органов виноградного растения и присущими им физиологическими процессами. В зависимости от сорта, климатических, метеорологических условий года и применяемой агротехники сроки прохождения фаз изменяются, но их последовательность сохраняется, что является биологической особенностью виноградного растения. Вегетация винограда начинается весной, с сокодвижения и распускания почек, когда устанавливается теплая погода со среднесуточной температурой воздуха 10°C. Эту температуру для виноградного растения принимают за биологический нуль. Заканчивается период вегетации осенью с наступлением похолодания и снижения среднесуточной температуры до 10°C и ниже. В зависимости от биологических особенностей культивируемых видов и сортов винограда, сорта — подвоя, а также от прогревания корнеобитаемого слоя почвы биологический нуль может несколько смещаться в сторону понижения или повышения. 1-я фаза — сокодвижение («плач» винограда)Наступает весной до набухания и в период набухания глазков, когда из всех порезов плодовых побегов и многолетних частей (рукавов) куста обильно вытекает прозрачная жидкость — пасока, что указывает на начало активной жизнедеятельности мочек корней, которые интенсивно всасывают из почвы воду с растворенными в ней питательными элементами и подают ее под давлением в надземную часть куста. Основное русло движения пасоки — флоэма. Поскольку в момент начала сокодвижения корневые волоски еще отсутствуют, следует считать, что вода может поглощаться через поверхность старых корешков, большинство из которых сохраняется в течение зимы и весной начинает жизнедеятельность. Наступление сокодвижения и степень его проявления зависят температуры, влажности почвы и состояния растения, сорта, подвоя и др. У сортов европейско-азиатского винограда сокодвижение начинается при прогревании почвы на глубине залегания основной массы корней до 7–9°C, у амурского и некоторых Американских видов (В. лабруска) — при 4,5–5,2°C, у В. вульпина — при 7–8°C, а иногда и при 4–6°C. В южных районах эта фаза начинается раньше, в северных несколько позже. У кустов, корневая система которых расположена ближе к поверхности почвы, сокодвижение наступает раньше. Более рослые кусты выделяют больше пасоки, чем слаборослые, что объясняется мощностью поглощающей части корневой системы. При похолодании и недостатке влаги в почве сокодвижение проходит слабее или прекращается, а при засушливой весне и повреждении корней морозами его не бывает; При благоприятных условиях с одного взрослого куста в сутки может вытечь до 2 л и более пасоки, однако при нормальной водообеспеченности это практически не истощает куста, поскольку пасока представляет собой почти чистую воду. Ее плотность равны 1,0007 г/см3. В 1л пасоки содержится до 2% сухих веществ, 2/3 из которых приходится на органические (сахара, аминокислоты) и 73 на минеральные (калий, кальций, фосфорная кислота, магний). Наиболее сильное выделение пасоки наблюдается при весеннем обновлении срезов побегов. Количество выделяемой пасоки при обновлении срезов составляет в Крыму 1,4 л, Одессе — 3, Краснодаре— 5, без обновления срезов — соответственно 0,2, 1,4 и 2 л. При орошении объем выделяемой пасоки увеличивается. В начале фазы сокодвижения пасоки выделяется мало, затем количество ее увеличивается и к концу фазы уменьшается. Наиболее активное сокодвижение продолжается не более 10–15 дней, а общая продолжительность его в зависимости от района и условий года составляет от 2 до 66 дней. Пасока имеет слабокислую реакцию (рН 6,8). Рукава и побеги становятся упругими, гибкими поэтому их легко подвязывать к опоре и укладывать отводки. В этой фазе проводят также прививку врасщеп. На молодых привитых виноградниках удаляют поверхностные корни и подвойную поросль. На участках осуществляют плантаж, устанавливают и ремонтируют шпалеры. В районах неукрывного виноградарства в фазе сокодвижения продолжают обрезку кустов. В районах укрывного виноградарства этим работам предшествует открытие кустов. С распусканием почек и развитием на побегах листьев, которые начинают испарять воду, сокодвижение прекращается. 2-я фаза — рост побегов и соцветий.С установлением устойчивой температуры воздуха выше 10°C и поступлением из корней в надземную часть воды с растворенными в ней питательными элементами начинает активизироваться жизнедеятельность всех органов растения: усиливается их дыхание, увеличивается тургор клеток, возрастает активность ферментов, с помощью которых осуществляется перевод нерастворимых органических соединений (крахмала) в растворимые (сахара). К точкам роста поступают питательные вещества, происходит интенсивное деление клеток. Глазки начинают набухать. В это время наиболее активно растут верхушка и междоузлия эмбрионального побега, чтоуриводит к распусканию глазка. Его покровные чешуи раскрываются, в густом войлоке появляются разрывы, из которых показывается верхушка растущего побега с первыми ярко-зелеными листочками. Чем быстрее повышается температура, тем дружнее распускаются почки. Первыми на побеге в связи с проявлением полярности распускаются почки верхних глазков, затем нижерасположенных. Для начала распускания почек необходима сумма активных температур (выше 10°C) в пределах 145–258°C. При более высоких температурах продолжительность этого периода уменьшается, при более низких — увеличивается. Оптимальная: температура воздуха в этой фазе 20–30°C, влажность почвы — 80–85% наименьшей влагоемкости (НВ). В южных районах виноградарства почки начинают распускаться в конце марта — первой половине апреля, в более северных— в начале мая. Набухание глазков и распускание почек — ответственный, критический период в малом годичном цикле виноградного растения. В это время проходит процесс дифференциации соцветий в почках глазка, идет образование в них осей 2–и 3–го порядков. При нарушении питания зачатки соцветий слабо дифференцируются и образуют усиковые недоразвитые соцветия или усик. Если процесс распускания почек глазка под влиянием высоких температур проходит ускоренными темпами, в почках глазка не успевают полностью пройти процессы дифференциации зачаточных соцветий, в результате чего снижается количество нормально развившихся соцветий, повышается количество недоразвитых соцветий или усиков. И, наоборот, в случае распускания почек глазка в течение более продолжительного периода улучшается развитие в них генеративных органов. В этой фазе вместе с ростом побегов продолжаются закладка и формирование скороспелых пазушных и зимующих почек, усиленное ветвление корней, образование новых корешков (мочек. и корневых волосков), начинается активная жизнедеятельность всех органов растения. При этом рост надземной и корневой систем проходит синхронно. Для их хорошего роста и развития очень важно, чтобы осенью и ранней весной растение было обеспечено Достаточным запасом питательных элементов и влаги, что достигается соответствующей агротехникой. В самом начале фазы распускание почек и рост побегов идут за счет осенних запасов органических веществ, накопленных в корнях и стеблях растения в предшествующем году, в связи с чем в этот период значительно снижаются запасы углеводов в корнях и побегах. С развитием листьев продукты фотосинтеза расходуются в основном на построение новых тканей растущих органов куста. В начале фазы побеги растут медленно, затем их рост постепенно усиливается (до 7—10 см в сутки) и в конце фазы снова затухает. В период наиболее интенсивного роста побегов на растении образуется до 80% листьев, большинство из которых достигает своей максимальной величины. Рост побегов в зависимости от особенностей сорта и условий внешней среды продолжается, как правило, до начала вызревания побегов, а иногда и до физиологической зрелости ягод. Общая лродолжительность фазы — 35–55 дней. Она зависит от метеорологических условий, в первую очередь от температуры воздуха. В конце фазы, к началу цветения, побеги достигают 60% полной длины. Вершина кривой роста побегов приходится на предшествующий цветению период, цветение или на период сразу после него. Это зависит от температуры воздуха. Интенсивность роста побегов в дневные часы (12–16) выше, чем в ночные. Однако это соотношение изменяется в зависимости от периода роста побегов. В начальные этапы, когда ночные температуры ниже дневных, прирост побегов идет более активно днем. В конце фазы эта закономерность может изменяться в пользу ночного периода. Следовательно, в данном случае большое значение имеет не столько световой, сколько температурный фактор. Продолжительность интенсивного роста побегов не зависит от биологических особенностей сорта, а обусловлена метеорологическими условиями, в первую очередь температурой. Оптимальная температура для роста побегов 25–30°C. На интенсивности роста побегов сказывается и близость их к корневой системе: чем ближе расположен побег к корневой системе, тем интенсивнее он растет. Темпы роста побегов у растений, сформированных на высоком штамбе, несколько ниже. В силу полярности развившиеся в верхней и нижней частях плодового побега плодоносные и бесплодные побеги растут более интенсивно. У растений, привитых на сильнорослых подвоях, побеги привоя растут быстрее. Рост пасынков зависит от их расположения на побеге. Наиболее энергично пасынки растут на 3–5–м узлах от основания развившихся побегов. В этой фазе полностью формируются соцветия, а в пазухах листьев по всей длине побега закладываются пазушные скороспелые (пасынковые) и зимующие почки. На нижних узлах побега почки менее развиты из-за недостатка питательных веществ, так как их формирование приходится на начало фазы, когда на побеге еще мало листьев, которые не достигли нормальной величины, поэтому характеризуются слабой фотосинтетической активностью. Наиболее развитые почки формируются в период усиленного роста побегов в их средней части. В это время листья достигают полного развития, содержат много хлорофилла и отличаются активной фотосинтетической деятельностью. В фазе роста побегов и соцветий необходимо следить за погодными условиями. На случай снижения температуры воздуха до 0–1°C проводят агротехнические мероприятия по защите распускающихся почек и растущих молодых побегов от повреждения поздними весенними заморозками. В начале фазы, когда видны соцветия, обламывают побеги, ненужные для плодоношения и формирования кустов, а при достижении побегами 25–30 см длины их подвязывают к опоре. Начинают пасынкование и применяют профилактические обработки виноградников пестицидами против милдью и оидиума, ведут борьбу с вредителями. Перед цветением прищипывают побеги и подкармливают растения. Периодически рыхлят почву для уничтожения корки и сорняков. На маточниках подвоев проводят пасынкование, подвязку побегов, удаление соцветий. В течение этой фазы можно уже ориентировочно определить величину урожая текущего года. 3-я фаза — цветение.Начинается раскрыванием и сбрасыванием венчика (колпачка) с бутона цветка. Первыми распускаются центральные бутоны и расположенные на осях периферийных соцветий ближе к основанию побега (что объясняется лучшим их прогреванием), затем боковые и вышерасположенные бутоны. На кустах с низким штамбом цветение наступает раньше, чем с высоким, и у крупных соцветий раньше, чем у мелких. Начало цветения винограда зависит главным образом от температуры. Цветение начинается при температуре воздуха 16°C, а оптимальной для этого процесса является температура 20–30°C. При температуре ниже 11°C пыльца не прорастает, а при 11–15°С она прорастает медленно. При более высокой температуре и относительной влажности ниже 45% снижается завязываемость ягод из-за быстрого высыхания секреторной жидкости на рыльце. На ход цветения сильно влияет температура предшествующего дня. При снижении температуры воздуха до 11–15°C, затяжных дождях и туманах венчики плохо сбрасываются, а пыльца в дождливую погоду смывается с рыльца. При благоприятных для цветения погодных условиях рыльце пестика цветка сохраняет восприимчивость к оплодотворению в течение четырех — шести дней после раскрытия бутонов. Бутоны в соцветиях начинают распускаться в 6–8 ч утра. Количество раскрывшихся бутонов увеличивается до 9—10 ч утра. К 11 ч распускание бутонов приостанавливается и только во второй половине дня (с 15 до 16 ч) начинается вторая волна цветения, однако она менее интенсивна и непродолжительна. В благоприятных погодных условиях продолжительность цветения одного соцветия составляет 4–8, сорта — 8-14 дней. За начало цветения принимают календарный день, когда раскроется 5% цветков, массовое (разгар)— 60–70 и конец цветения — 25–30%. Цветение и оплодотворение (рис. 29) цветков винограда происходят следующим образом. В период готовности цветка к цветению в месте соединения венчика с диском образуется разделяющий слой ткани. В день цветения вследствие повышения температуры и снижения влажности воздуха, а также за счет различия важности наружных и внутренних лепестков венчика начинается их постепенный обрыв, и венчик спадает в виде колпачка. Пыльники на тычинках растрескиваются, и из них высыпается пыльца. Попадая на рыльце пестика, где к этому времени появляются капельки секреторной жидкости, пыльца прилипает к нему и при благоприятных условиях— оптимальной температуре 25–30°C и влажности воздуха не менее 75–80% — прорастает в многочисленные пыльцевые трубки, которые проходят по грифельному слою столбика пестика, проникают через пыльцевход в зародышевый мешок, семяпочку, и спермий из пыльцевой трубки оплодотворяет яйцеклетку. Из каждой оплодотворенной семяпочки развивается одно семя, а из оплодотворенной яйцеклетки — зародыш. В зависимости от количества оплодотворенных семяпочек в ягоде может развиваться от одного до четырех семян и более. После оплодотворения хотя бы одной семяпочки начинается рост завязи. Она увеличивается в размере, столбик и рыльце отсыхают, и образуется ягода. Исследованиями установлено, что в зародышевый мешок проникает много пыльцевых трубок. Однако благодаря избирательной способности яйцеклетки с ней сливается содержимое только одной пыльцевой трубки, наиболее соответствующее развитию устойчивого организма. Содержимое других пыльцевых трубок служит для питания развившегося зародыша. Для своего роста пыльцевая трубка расходует более 40% питательных веществ проводящих тканей коленхимного происхождения. Процесс оплодотворения длится сутки. Однако не все цветки в соцветиях оплодотворяются, а если и оплодотворяются, то в зависимости от сорта винограда происходит естественное осыпание 40–80% цветков и завязей, что считается нормальным явлением. Максимальное осыпание завязей наблюдается на 9—15–й день после массового цветения винограда. Выявлено, что после оплодотворения в ягоды завязываются не более 15–20% цветков соцветиях. Важно, чтобы после осыпания цветков и завязей в соцветии осталось такое количество завязей, которое соответствует получению грозди с нормальной плотностью ягод, характерной для конкретного сорта. Но бывают случаи, когда наблюдается чрезмерное осыпание завязей, в результате образуются рыхлые грозди с малым количеством ягод, что резко снижает величину урожая Низкая завязываемость ягод зависит от ряда причин, прежде всего от плохого опыления и оплодотворения цветков, что, в свою очередь, обусловливается неблагоприятными для цветения погодными условиями, недостатком питательных элементов и воды, дефектами в строении цветка, отсутствием соответствующего опылителя, особенно для сортов с функционально-женским типом цветка, а также биологической особенностью некоторых сортов, их склонностью к осыпанию цветков (Мускат гамбургский, Мускат венгерский, Мускат александрийский, Саперави, Сенсо). В каждом конкретном случае выясняют причины чрезмерного осыпания завязей и принимают меры к их устранению: дополнительное и искусственное опыление сортов, имеющих как обоеполый тип цветка, так и функционально-женский, прищипывание верхушек побегов перед цветением и их кольцевание, высокий фон агротехники и др.
Рис. 29. Оплодотворение винограда: пыльцевая трубка (1), проникшая в зародышевый мешок (2); семяпочка (3) (по Баранову). Цветение винограда во всех районах наблюдается в июне, а в некоторых районах республик Средней Азии — во 2–3–й декаде мая. У разных сортов винограда сроки цветения могут не совпадать. Поэтому изучение фазы цветения имеет большое значение при подборе опылителей. В период цветения на виноградниках продолжают обработку почвы, опыливание против оидиума и опрыскивание против милдью, обработку насаждений бессемянных сортов гиббереллином. Однако в это время не рекомендуется проводить орошение, которое повышает влажность воздуха и снижает его температуру, что отрицательно сказывается на оплодотворении цветков. 4-я фаза — рост ягодПосле оплодотворения семяпочек у семенных сортов винограда развивается зародыш, формируется эндосперм и оболочка семени. Завязь увеличивается в размере и превращается в ягоду. Чем больше семяпочек оплодотворилось в завязи, тем быстрее развивается околоплодник и крупнее ягода. В росте ягод наблюдается 3 этапа. Первый этап — интенсивного роста — следует сразу за процессами опыления и оплодотворения. Затем наступает второй этап, когда рост приостанавливается (перед началом созревания ягод), и, наконец, третий этап — новая волна роста (в период их созревания). Эти изменения обусловливаются специфическими внутренними причинами, характерными для каждого этапа. Первый этап роста ягод проходит путем деления клеток под воздействием физиологически активных веществ (ФАВ). За относительно короткий промежуток времени ягоды набирают 60–70% своей массы. Этот процесс протекает синхронно с ростом семян и приостанавливается с прекращением их роста, так как молодые растущие семена являются центрами продуцирования гиббереллинов и ауксинов, которые стимулируют рост околоплодника ягоды. В связи с этим на интенсивность роста семенных сортов винограда в значительной степени влияет количество семян в ягоде, которое определяется особенностью строения завязи, числом гнезд (от 1 до 4–5) и семяпочек в них (от 1 до 7). Но, как правило, более 95% завязей двухгнездные и в каждом гнезде по 2 семяпочки. С увеличением числа семян в ягоде на одно масса мякоти возрастает примерно на 10%, при наличии четырех семян в ягоде — на 35–40%. От успешного процесса опыления и оплодотворения в значительной степени зависит величина урожая. У бессемянных сортов рост ягоды происходит под влиянием ростовых веществ, образовавшихся в процессе опыления и начала роста и развития рудиментов семян. С окончанием роста семян и началом их созревания первая волна роста ягод приостанавливается, и в них происходят качественные изменения, направленные на начало созревания ягод. Пока ягода растет, она остается зеленой, на ее поверхности имеются устьица и она ассимилирует. Однако вырабатываемые ягодой в этот этап продукты фотосинтеза компенсируют лишь 1/5 потребности, остальные органические вещества поступают на рост и развитие ягоды из листьев и запасов, сосредоточенных в многолетних частях стебля и корней. Во второй этап, когда ягоды достигнут величины горошины, устьица на кожице деформируются, и превращаются в чечевички, при этом резко снижается ассимиляция углерода. В ягодах уменьшается содержание хлорофилла, и крахмала. В начале созревания ягод наступает третий этап их роста, проходящий за счет растяжения клеток мякоти и заполнения их соком. Ягоды достигают характерной для сорта величины, стенки их клеток растягиваются и делаются тонкими. К этому времени содержание кислот в ягодах максимальное (20–30 г на 1 кг их массы), а сахара только начинают накапливаться (0,5–0,6%). В фазу роста ягод листья нижнего и среднего ярусов полностью заканчивают рост, интенсивность их фотосинтеза достигает максимальной величины, побеги, хотя и замедленно, но продолжают рост в длину и особенно в толщину, в пазухах листьев продолжают закладываться зимующие почки и в них бугорки соцветий для обеспечения урожая в будущем году, идет интенсивное ветвление корней. В клетках всех органов уменьшается содержание воды и происходит накопление крахмала, начинается вызревание побега от его основания к вершине. Интенсивность прохождения этой фазы в значительной степени зависит от температуры воздуха. Наиболее сильный рост ягод наблюдается при температуре 25–30°C. Сумма активных температур от начала цветения до начала созревания ягод составляет 800–880°C. В зависимости от сорта, метеорологических условий и места произрастания фаза роста ягод длится 30–60 дней, а иногда и более. Растение при этом особенно нуждается в питательных элементах, влаге и хорошем освещении. В этой фазе на виноградниках проводят орошение, подкормки, продолжают подвязывать побеги к опоре и пасынковать их. Ведут борьбу с вредителями и болезнями, культивируют почву в междурядьях и рыхлят в рядах. В насаждениях сортов, у которых приостановился рост побегов, приступают к чеканке — удалению верхушек побегов. 5-я фаза — созревание ягодВнешне выражается в изменении цвета ягоды. Она теряет травянистую зеленую окраску вследствие разрушения хлорофилла. У белых сортов ягода становится матово-белой со слабыми признаками прозрачности. У сортов с окрашенной ягодой на кожице появляются темные пятна. Ягода делается мягкой, эластичной, на ее поверхности появляется восковидный налет (пруин), хорошо заметный у темноокрашенных сортов, предохраняющий ягоду от гниения, консистенция ее разжижается. Ягода достигает свойственной сорту величины за счет растяжения клеток и заполнения их соком. Существенно изменяется химический состав сока ягод. В начальный этап роста ягоды все. питательные вещества затрачиваются на ее рост. Незадолго до начала созревания в ягодах заметно увеличивается сахаристость сока, однако поворотный момент наступает позже, в начале созревания ягоды, когда скачкообразно возрастает содержание сахаров и начинает снижаться кислотность. Интенсивность накопления сахаров в ягоде превышает интенсивность роста ягоды. В течение недели количество сахаров в созревающих ягодах увеличивается в 6–7 раз. В (соке созревающих ягод винограда сахара представлены в основном глюкозой, фруктозой и сахарозой. В начальный этап созревания ягод глюкоза превалирует над фруктозой, затем их соотношение уравновешивается, ко времени полной зрелости ягод фруктозы бывает несколько больше, чем глюкозы, на имеются и сортовые различия. В начале созревания в ягодах преобладает яблочная и в небольшом количестве содержится винная кислота. По мере созревания количество винной кислоты значительно увеличивается и Достигает 80–90% от содержания всех кислот в ягоде. Однако У отдельных сортов количество винной и яблочной кислот находится в равных соотношениях. В процессе созревания в ягодах повышается содержание азотистых, красящих и уменьшается количество дубильных веществ. В созревающей ягоде отсутствует крахмал, но в начале фазы его много в гребнях и ножке ягоды, к концу фазы он исчезает. У ягод винограда различают 2 формы зрелости: физиологическую, (полную) зрелость ягод и техническую — товарную. Последняя определяется кондициями сахаристости и кислотности сока ягод, соответствующими направлению использования винограда. При полной физиологической зрелости ягоды винограда приобретают характерную для сорта массу, окраску, вкус и аромат. Содержание сахара в ягоде максимальное. Семена к этому времени достигают своей нормальной величины, становятся твердыми и приобретают коричневую окраску. При наступлении полной зрелости ягод абсолютное увеличение содержания в них сахаров прекращается, но относительное, вследствие испарения воды, продолжается. В этом случае ягоды начинают перезревать, заизюмливаться, завяливаться. У перезревших ягод с белой окраской на кожице появляется золотисто-желтый загар с темно-коричневыми пятнами. Если созревшие ягоды вовремя не собрать, они теряют упругость, сморщиваются, уменьшаются в объеме, вследствие чего урожайность виноградных насаждений снижается. С момента, полной зрелости ягод приток питательных веществ к ним прекращается. Вырабатываемые листьями органические вещества идут на вызревание побегов и откладываются в запас в многолетних частях куста (штамбе и рукавах) и корнях. Побеги и пасынки почти прекращают рост и идет их вызревание. Полностью завершается формирование глазков, в них происходит дифференциация 1–го и образование бугорков 2–и 3–го соцветий плодоносных почек. Продолжительность фазы созревания ягод у ранних сортов 20–30, у поздних — 50–60 дней. Чем выше температура воздуха и обеспеченность растений водой, тем быстрее созревают ягоды и в них накапливается больше сахаров. Дожди и повышенная влажность почвы вызывают растрескивание ягод и затягивают их созревание. Оптимальная температура для созревания ягод 28–32°C. При температуре ниже 14–16°C и выше 40°C ягоды созревают медленно. Сумма активных температур от начала созревания до физиологической зрелости ягод составляет 1048–1644°C. Вся агротехника на виноградниках в этой фазе должна быть направлена на создание условий, наиболее благоприятных для созревания ягод и накопления в них сахаров в соответствии с кондициями для разного использования урожая. Ведут строгий контроль за химическим составом ягод. В случае необходимости продолжают опыливание виноградников против оидиума и опрыскивание против милдью, но пестициды применяют с большой осторожностью, прекращают поливы, после выпадения дождей проводят рыхление почвы, продолжают чеканку побегов. Осуществляют апробацию, массовую и клоновую селекцию, предварительное определение величины ожидаемого урожая. В зависимости от наступления физиологической зрелости ягод сорта винограда условно делят по срокам созревания: на очень ранние, ранние, раннесредние, средние, среднепоздние, поздние и очень поздние. Для каждой из этих групп требуется определенное количество дней и определенная сумма активных температур. 6-я фаза — листопадВ этой фазе затухает или прекращается вегетативный рост побегов и происходит интенсивное их вызревание. Нижние междоузлия побегов (с 1–го по 7–е) вызревают медленно, в течение 10–15 дней, вышерасположенные (с 8–го по 14–е) быстрее, за 4–5 дней. Нормальному ходу вызревания побегов в значительной степени способствует снижение температуры воздуха до 15–10°C, а также резкая амплитуда колебаний дневной и ночной температур, характерная для осеннего периода, укороченный световой день и наступление заморозков. Интенсивность и сроки вызревания побегов не зависят от принадлежности винограда к группам по срокам созревания ягод, а связаны с силой роста побегов. Начало вызревания побегов не совпадает с началом созревания ягод. У амурского винограда и его гибридов побеги начинают вызревать раньше, чем ягоды, а у некоторых среднеазиатских сортов ягоды созревают значительно раньше, чем начинается вызревание побегов. При вызревании в побегах происходят сложные морфологические, анатомические и биохимические процессы, связанные с накоплением крахмала и превращением его в сахара, изменением структуры белковых веществ (что предохраняет их от свертывания при замерзании клеток) и коллоидов цитоплазмы, уменьшением количества свободной и связанной воды, повышением концентрации клеточного сока, утолщением и одревеснением ксилемы, флоэмы и сердцевинных лучей. В результате накопления в них лигнина, гемицеллюлозы и минеральных веществ идет образование пробкового камбия. Наружные слои коры высыхают и образуется корка. Побег приобретает желтовато-коричневую окраску. Все эти биолого-физиологические процессы способствуют закалке побегов и повышению их зимостойкости, благодаря чему глазки вызревших побегов в зависимости от сортовых особенностей растений выдерживают морозы до —16–25°C, а сами побеги до —20–27°C. Черенки, заготавливаемые с хорошо вызревших побегов, обеспечивают получение высококачественного посадочного материала. Такие побеги обладают более высокой морозоустойчивостью, без повреждений переносят зиму, что обеспечивает сильный рост куста и его высокую урожайность в следующем году. В южных районах виноградарства к концу вегетации побеги вызревают почти на всю длину своего прироста. В районах укрывной культуры из-за наступления ранних осенних заморозков побеги успевают вызревать на 65–70% прироста. Поврежденные заморозками невызревшие верхушки побегов и листья высыхают и опадают. В этих районах виноградники укрывают, обычно не дожидаясь листопада. Если же заморозков не бывает, то наступившее похолодание, характерное для осеннего периода, вызывает прекращение фотосинтеза в листьях и в них идут процессы гидролиза и оттока питательных веществ. В результате разрушения хлоропластов в листьях образуются красящие пигменты, придающие им характерную осеннюю окраску. У белых сортов листья становятся желтыми, у темноокрашенных пурпурно-красными с различными оттенками. У основания черешка листа в месте прикрепления его к побегу образуется плотная пробковая отделительная ткань, и начинается естественный листопад. Период от сбора урожая до вызревания побегов и листопада принято называть резервной фазой. В это время заканчивают сбор урожая винограда, проводят глубокое рыхление почвы с одновременным внесением органических и минеральных удобрений, а также предварительную обрезку кустов и заготовку черенков. В укрывных зонах виноградарства кусты укрывают на зиму. Вызреванием побегов, листопадом и подготовкой растения к зимовке (закалкой) заканчивается последняя фаза вегетации, после чего растение вступает в период относительного покоя. Фазы вегетации у сортов винограда изучают при помощи фенологических наблюдений, что позволяет установить календарные сроки появления у растений определенных морфологических изменений, характеризующих начало и конец каждой фазы. Для этого на типичных для хозяйства участках насаждений винограда каждого сорта выделяют небольшую делянку или отмечают этикетками и краской отдельные кусты в количестве не менее 25–30 шт., за каждым из которых или за всеми вместе ведут наблюдения. За начало фазы принимают календарную дату, при которой признаки фазы будут отмечены примерно у 5% кустов; массовое вступление в фазу — при появлении признаков фазы у 50–60% кустов и окончание фазы отмечают в тот день, когда примерно у 25% растений признаки фазы будут утрачены. Показатели записывают в журнал фенологических наблюдений. Способность виноградного растения переходить из состояния вегетации к периоду покоя — характерная биологическая особенность, которая сохраняется при возделывании винограда как в открытом, так и защищенном грунте (теплицах, оранжереях) и в различных экологических условиях — тропиках, субтропиках и т. д. При этом сдвигаются в основном лишь календарные сроки прохождения растениями фаз вегетации. Происходящие в годичном цикле развития растения морфологические изменения и физиологические процессы обусловлены сменой времен года. Продолжительность периода вегетации зависит как от сорта, района выращивания винограда (табл. 1), так и от метеорологических условий года. У сортов очень раннего срока созревания период вегетации короткий — 95—120 дней, у сортов позднего срока созревания он длится более 170 дней. С продвижением культуры винограда в северные районы продолжительность периода вегетации уменьшается, что объясняется компенсацией интенсивности освещения, увеличенной долготой дня и достаточно высокими температурами летом. В случае продвижения винограда в горы период вегетации также укорачивается, что обусловливается понижением температуры воздуха и почвы. В районах неукрывного виноградарства длительность периода вегетации определяют по числу дней от начала сокодвижения до естественного листопада, укрывного виноградарства — от начала распускания почек до полной зрелости ягод. При изучении фаз вегетации винограда фиксируют среднесуточную температуру воздуха и количество выпадающих осадков. 1. Результаты фенологических наблюдений за растениями сорта Ркацители в различных районах его выращивания (средние данные за 5—10 лет)
В период вегетации следят за динамикой роста побегов, их вызреванием и созреванием ягод, что позволяет выявить закономерности роста и развития и реакцию растений на изменения условий внешней среды. Динамику роста побегов определяют путем измерения длины (см) одних и тех же нормально развивающихся побегов через каждые 10–20 дней, динамику вызревания побегов — непосредственно на кустах по наружной окраске их корки. Для этого через каждые 4–5 дней одни и те же побеги осматривают и измеряют длину их вызревшей части (или подсчитывают количество вызревших междоузлий) и выражают в процентах к общей длине побега. Степень зрелости ягод винограда определяют по количественному накоплению в соке сахаров и кислоты в процессе созревания, сначала через каждые 3–5 дней, а при приближении к требуемой кондиции — ежедневно. Многолетние (трех — пятилетние) фенологические наблюдения с Учетом суммы активных температур за каждую фазу вегетации позволяют правильно определить сортимент, соответствующий конкретным почвенно-климатическим районам промышленной культуры винограда. На основе материалов многолетних фенологических наблюдений в хозяйствах составляют технологические карты Ухода за виноградными насаждениями и сбора урожая. |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||