|
||||
|
|
Часть I. Гематология. Общая часть Глава 1. Кроветворение Кроветворение – сложный процесс, включающий в себя много стадий клеточных дифференцировок, итогом которых является выход в кровеносное русло таких форменных элементов, как лейкоциты, эритроциты и тромбоциты. Основная функция эритроцитов заключается в транспорте кислорода, углекислого газа, белков, углеводов, аминокислот, гормонов, ферментов, микроэлементов в органы и ткани. Они принимают участие в процессе остановки кровотечения, в формировании иммунитета, тем самым осуществляя защитную функцию. Лейкоциты вырабатывают антитела, которые разрушают вредные антигены, попавшие в организм человека, тем самым принимая участие в иммунных реакциях. Наиболее значимой функцией тромбоцитов является обеспечение остановки кровотечения путем образования тромбоцитарной пробки. Количество эритроцитов в крови взрослого человека составляет примерно 25 ? 1012/л, количество лейкоцитов – 3 ? 109/л, тромбоцитов – 15 ? 109/л. Объем форменных элементов крови составляет около 40% от всего объема крови, а остальные 60% приходятся на плазму, т. е. на жидкую часть крови. При отсутствии патологии продолжительность жизни эритроцитов в системном кровотоке составляет около 100–120 дней. Длительность циркуляции нейтрофила в кровеносном русле выражается временем полувыведения радиоактивной метки и составляет в среднем от 4 до 10 ч. По прошествии указанного временного промежутка нейтрофил поступает в ткани организма, где продолжительность его жизни также исчисляется несколькими часами. Период полувыведения радиоактивной метки моноцита составляет примерно 72 ч. После этого данный форменный элемент крови переходит в ткани. Там моноцит имеет возможность превратиться в фиксированный либо в блуждающий макрофаг, сохраняющий способность к делению. Указанное свойство является отличительной особенностью моноцитов: зрелые гранулоциты не способны к делению. Продолжительность жизни тканевого макрофага остается невыясненной. Длительность пребывания эозинофилов в кровотоке составляет около 5 ч, после чего они, так же как и вышеуказанные форменные элементы крови, мигрируют в ткани. Продолжительность нахождения в кровеносном русле таких форменных элементов, как базофилы, не установлена. Лимфоциты являются неоднородной группой кровяных клеток: они подразделяются на Т-лимфоциты и В-лимфоциты. При этом продолжительность жизни различных лимфоцитов неодинакова: одни клетки живут часы, другие – несколько лет. Длительность циркуляции в крови тромбоцитов – 8–9 дней. Различные форменные элементы крови могут различаться по степени своей зрелости. В различных условиях из органов, принимающих участие в процессе кроветворения, могут выходить как вполне зрелые, так и только еще созревающие, но уже осуществляющие свою первостепенную задачу клетки. Они могут фагоцитировать (поглощать и переваривать) чужеродные частицы, участвовать в переносе кислорода, формировать первичный тромб. Также в системном кровотоке возможно присутствие совсем незрелых клеточных элементов. Такими структурами могут являться предшественники эритропоэза, которые по своим морфологическим характеристикам ничем не отличаются от лимфоцитов. Необходимо помнить о том, что предшественники всех возможных ростков кроветворения обладают внешними признаками, полностью идентичными таковым у лимфоцита. В этой связи различать данные клетки по внешним морфологическим критериям не представляется возможным. Созревание форменных элементов в каждом из ростков кроветворения повторяет филогенез кроветворения. В процессе эволюции ядросодержащие эритроциты птиц и рыб трансформировались в безъядерные эритроциты млекопитающих. При этом созревание клеток красного ряда у человека включает в себя стадии ядросодержащих эритрокариоцитов в костном мозге, что повторяет их строение у рыб и птиц, а в кровоток выходят зрелые безъядренные эритроциты. Из вышеуказанного следует, что эмбриогенез крови в отличие от остальных органов и систем происходит непрерывно в течение всей жизни человека. Стволовые клетки Изучение процесса кроветворения проходило в направлении от уже дифференцированных зрелых форменных элементов крови к их предшественникам. Стадии, которые различные по своему строению кровяные клетки проходят в процессе своей дифференцировки, можно проследить на регенерирующем кроветворении. Такой процесс наблюдают после опустошения кроветворения посредством воздействия цитостатиков либо при иммунном агранулоцитозе (отсутствии в крови агранулоцитов). В конце дифференцировки (созревания) клетки-предшественницы трансформируются в зрелые элементы крови. В 1961 г. иностранными учеными-гематологами был разработан метод селезеночных колоний. Он основывался на том, что после трансплантации смертельно облученным мышам донорского костного мозга отмечается появление в их селезенке очагов кроветворных клеток, видимых невооруженным глазом. Применив хромосомные маркеры, представляющие собой стабильно измененные после облучения хромосомы, установили, что всякая подобная колония представляет собой клон, т. е. потомство всего лишь одной клетки, являющейся колониеобразующей единицей селезенки – КОЕс. При этом образование колонии из такого типа клетки происходит путем продукции миллионов дифференцированных клеток-потомков. Применяя методику селезеночных колоний, а также комбинируя ее с методом радиационных маркеров, установили, что на поверхности лимфоцитов располагаются те же маркеры, что на поверхности кроветворных клеток селезеночной колонии. Итогом данных исследований было доказано существование в костном мозге организма полипотентной клетки, являющейся общим предшественником всех возможных ростков кроветворения. Данная клетка получила название стволовой. При трансплантации участка костного мозга от здоровой мыши к смертельно облученной стволовая клетка может стать предшественницей всех форменных элементов крови. Впоследствии от данной мыши-реципиента ткань костного мозга можно трансплантировать другим мышам, получившим смертельную дозу облучения. В результате таких экспериментов сформировалось ошибочное мнение о том, что КОЕс – родоначальная стволовая клетка, имеющая морфологическую структуру большого лимфоцита и, с одной стороны, обладает способностью к практически абсолютному самоподдержанию, а с другой – к дифференцировке во всех возможных направлениях гемопоэза. Последующие эксперименты по трансплантации костного мозга от здоровой к облученной мыши установили, что КОЕс теряют имеющуюся у них первоначальную возможность повторений с каждой последующей трансплантацией. При этом было замечено, что через 2–3 пассажа у реципиента в составе колоний уже не возникают полипотентные стволовые клетки. Биологический смысл существования класса стволовых клеток заключается в способности их при возникновении определенных условий практически мгновенно переключать кроветворение на какое-либо одно из необходимых в данный момент времени направление, в чем также играют немаловажную роль механизмы регуляции. Полипотентные клетки-предшественницы Помимо колониеобразующих клеток селезенки, в культуре костного мозга присутствуют также клетки, обладающие высокой пролиферативной активностью. Такие клетки имеют способность дифференцироваться в направлении миелопоэза и лимфопоэза. Имеются сведения о наличии в организме особого вида клеток, располагающихся в классе стволовых выше КОЕс. Применяя довольно активные факторы стимуляции кроветворения, у мышей выявили клетки, образующие колонии из не подвергающихся дифференцировке бластных элементов. Данные полипотентные клетки, способные дифференцироваться по пяти различным направлениям, получили название колониеобразующей единицы гранулоцитарно-эритроцитарно-макрофагально-мегакариоцитарной (КОЕ-ГЭММ). В гематологической практике в настоящее время первой клеткой, относящейся к классу полипотентных клеток-предшественниц, является КОЕ-ГЭММ. Унипотентные клетки – предшественницы миелопоэза К данному классу клеток относятся те, которые способны к дифференцировке только лишь в одном направлении, т. е. дают начало одному виду форменных элементов крови. Предшественницей нейтрофилов, кроме гранулоцитарно-моноцитарной клетки-предшественницы, является независимая гранулоцитарная клетка-предшественница, обозначающаяся КОЕ-Г. Также существует самостоятельная клетка – предшественница эозинофилов. Унипотентная клетка – предшественница моноцитов (КОЕ-М) может иметь двоякое происхождение. Во-первых, она может являться потомком гранулоцитарно-моноцитарной клетки-предшественницы. Во-вторых, может быть потомством прочих полипотентных клеток, являясь при этом самостоятельной клеткой – предшественницей моноцитов. У человека имеется несколько типов клеток – предшественниц красного ряда (предшественники эритроцитов). К таковым относятся БОЕ-Э незрелая и БОЕ-Э зрелая. Это так называемые бурстобразующие единицы. Их характерной особенностью являются довольно крупные колонии. Кроме того, в организме человека присутствует и КОЕ-Э, т. е. эритроцитарная колониеобразующая единица – единица, являющаяся родоначальницей эритроцитарного ряда. Клетки-предшественницы красного ряда имеют ряд отличий друг от друга. В первую очередь они различаются местом локализации в кроветворных органах, а также местом циркуляции. Также данный класс клеток различен по размерам колоний, которые они образуют в результате культивирования на питательных средах, неодинаковому времени насыщения клеток гемоглобином, чувствительности к эритропоэтину и ряду других факторов регуляции, типу синтезируемого гемоглобина, доминированию в каком-либо определенном возрасте человека. БОЕ-Э незрелая (примитивная) относится к самым ранним клеткам-предшественницам, осуществляющим свое развитие исключительно в направлении красного ростка. Указанная клетка является неоднородной единицей. В культуре присутствует довольно ранняя БОЕ-Э – незрелая, образующая свою колонию, имеющую пик гемоглобинизации (насыщения гемоглобином) уже на 18–21-й день дифференцировки. Однако большую часть БОЕ-Э незрелых образуют клетки, дающие начало своим большим колониям, представляющим собой скопления более 16 единиц. В том случае, если количество клеток, образующих колонии первых БОЕ-Э незрелых, достигает нескольких десятков тысяч, то в другом – тысячи. Пик гемоглобинизации образующихся колоний БОЕ-Э незрелой приходится на 14-й день. Клетки БОЕ-Э незрелой отличаются от иных присутствием крупных молодых ядер. БОЕ-Э зрелая дает гораздо мелкие колонии. Наиболее высокий уровень гемоглобинизации данных колоний в культуре выявляется уже на 10–12-й день культивирования (гораздо раньше колоний БОЕ-Э незрелой). Также по сравнению с колониями БОЕ-Э незрелой колонии клетки, образующие колонии БОЕ-Э зрелых клеток-предшественниц, имеют меньшие размеры и более компактное ядро. КОЕ-Э – колониеобразующая эритроцитарная единица – относится к наиболее зрелым клеткам – предшественницам красного ряда, данные колонии образованы из 50–100 клеток. Наиболее высокий уровень гемоглобинизации в культуре данных колоний выявляется на 7-й день. Эритрокариоциты по сравнению с клетками бурст имеют более компактное ядро и небольшие размеры. Также клетки-предшественницы красного ряда различаются между собой по типу продуцируемого ими гемоглобина. Зрелые клетки-предшественницы – БОЕ-Э зрелая и КОЕ-Э – продуцируют гемоглобин взрослого типа (НЬА) и лишь 1–2% гемоглобина фетального типа (HbF). В то же время оказалось, что ранние клетки-предшественницы красного ряда – БОЕ-Э незрелые продуцируют колонии, содержащие в основном HbF. Большая часть БОЕ-Э незрелых дает менее 20% колоний, содержащих HbF, причем одни колонии содержат только HbF, другие – только НЬА, отдельные колонии (каждая происходит из одной клетки) содержат и HbF, и НЬА. В костном мозге взрослого человека превалируют зрелые бурсты, а также колониеобразующие эритроцитарные единицы (КОЕ-Э), именно они и созревают (пролиферируют) и дают зрелое потомство – эритроциты, содержащие гемоглобин взрослого типа – НЬА. Морфологически распознаваемые клетки Вслед за унипотентными клетками, характеризующимися, как и более ранние предшественники, морфологическим однообразием и отсутствием возможности их морфологической дифференцировки, в схеме кроветворения располагается класс делящихся и морфологически вполне дифференцируемых (распознаваемых) клеток. В ряду гранулоцитов от миелобласта до миелоцита все клетки проходят митотический цикл, т. е. их созревание связано с делением. Последняя делящаяся клетка гранулоцитов – миелоцит – после деления превращается в метамиелоцит, который без деления в результате уплотнения ядерного хроматина трансформируется в палочкоядерный и сегментоядерный гранулоциты. Образование моноцитов проходит хорошо определяемые при обычной окраске стадии дифференцировки: монобласт, промоноцит, моноцит. Следует отметить, что в отличие от гранулоцитов, где зрелые – палочкоядерные и сегментоядерные – элементы к дальнейшему делению не способны, зрелые моноциты делятся и могут превращаться в макрофаги. В макрофагальном ряду кроветворения выделяют макрофагальный бласт, промакрофаг, макрофаг. Различаемые при обычной окраске эритрокариоциты представлены эритробластом, пронормоцитом, нормоцитами: базофильным, полихроматофильным и оксифильным. Базофильный нормоцит окрашивается основными красителями, оксифильный – кислыми (низкое значение рН), полихроматофильный – красителями с любым значением рН. За исключением оксифильного нормоцита, все эритрокариоциты подвергаются процессу деления и дифференцировки. Следовательно, каждой последующей генерации клеток должно быть вдвое больше предыдущей. В схеме кроветворения, предложенной еще И. А. Кассирским и Г. А. Алексеевым, параллельно ряду нормальных эритрокариоцитов располагался мегалобластический ряд. Мегалобласты, располагающиеся в костном мозге плода, после рождения встречаются лишь при дефиците в организме витамина В12 или фолиевой кислоты. При этом кроветворение является патологическим – дефицит указанных витаминов ведет к нарушению синтеза ДНК и РНК в эритрокариоцитах. При нарушении синтеза ДНК и РНК, вызванном приемом некоторых лекарственных препаратов (например, метотрексата или цитозара), эритрокариоциты приобретают уродливое мегалобластическое строение. В схему нормального кроветворения мегалобластический ряд в настоящее время не входит. Между тем мегалобластический характер обнаруживают эритрокариоциты при некоторых анемиях, не связанных с дефицитом витамина В12 (при аутоиммунной гемолитической анемии), а также при некоторых формах миелоидных лейкозов – остром малопроцентном лейкозе, эритромиелозе (остром и хроническом) – клетки красного ряда тоже имеют мегалобластическое строение. Наконец, мегалобластический эритропоэз встречается при алкоголизме. Следовательно, кроветворение при В12 или фолиеводефицитных анемиях, как и при всех перечисленных состояниях, совершается не по беспорядочно анархическому варианту, как можно было бы ожидать при нарушениях синтеза ДНК и РНК в клетках, а по морфологически и функционально однотипному образцу с выраженной гиперхромией клеток (увеличение цветового показателя), иногда с высоким процентом клеток с гемоглобина F. Регуляция кроветворения Регуляция кроветворения неодинакова на разных его ступенях. Стволовые клетки и ранние клетки – предшественницы гемопоэза контролируются посредством близкодействующей регуляции, которая обеспечивается за счет непосредственного взаимодействия с соседними кроветворными клетками и клетками стромы костного мозга. Поздние клетки-предшественницы регулируются гуморальными факторами. Увеличение и разделение стволовых клеток находятся под воздействием как стромальных клеток (образующих строму органа), так и кроветворных клеток – ближайшего потомства стволовой клетки, – и клеток лимфатической и макрофагальной природы. При облучении костного мозга в дозах ниже 5 Гр в крови наблюдается абортивный подъем лейкоцитов, тромбоцитов, ретикулоцитов, который отодвигает окончательное восстановление состава периферической крови на более поздний срок по сравнению со сроками восстановления после облучения костного мозга в более высоких дозах. Очевидно, уцелевшие после облучения ранние клетки-предшественницы создают абортивный подъем показателей периферической крови, временно обеспечивают кроветворение и своим существованием задерживают появление кроветворения из стволовой клетки, которое приходит на смену абортивному. В регуляции размножения ранних полипотентных и унипотентных клеток-предшественниц немаловажное значение имеет их взаимодействие с Т-лимфоцитами и макрофагами. Данные клетки действуют на клетки-предшественницы с помощью продуцируемых ими факторов – веществ, содержащихся в мембране и отделяющихся от нее в виде пузырьков при тесном контакте с клетками-мишенями. Регуляция эритропоэза Из регуляторов ранних клеток – предшественниц красного ряда особый интерес представляет бурст-промоторная активность (БПА). БПА обнаруживается уже при печеночном кроветворении у плода, но в основном ее роль проявляется в эритропоэзе взрослого. Стимулирующим действием на БОЕ-Э незрелые колонии обладают преимущественно костномозговые макрофагальные элементы, используемые в культуре в низкой концентрации, тогда как высокая концентрация этих клеток ведет к препятствию размножения бурстобразующих единиц. Влияние моноцитарно-макрофагальных элементов на клетки красного ряда разнообразно. Так, макрофаги являются одним из основных экстраренальных (располагающихся вне почек) источников эритропоэтина. У плода эритропоэтин выделяют купферовские клетки печени. У взрослого купферовская клетка вновь начинает продуцировать эритропоэтин в условиях регенерирующей печени. Для красного ряда характерно постепенное нарастание чувствительности к эритропоэтину, основному гуморальному регулятору эритропоэза, от ранних клеток-предшественниц к поздним. Гипоксия – снижение кислорода в тканях – стимулирует выработку эритропоэтина. Постоянная или кратковременная гипоксия в эксперименте на мышах с имплантированной диффузионной камерой вела к повышенной пролиферации БОЕ-Э незрелых [Harigaya et al., 1981]. В то же время опыты с гипоксией у обезьян в гипобарической камере показали значительное повышение у них HbF-содержащих эритроцитов в крови. Гипоксия является следствием снижения уровня кислорода во внешней среде (при подъеме на большую высоту), дыхательной недостаточности при поражении легочной ткани, повышенного потребления кислорода (например, при тиреотоксикозе). Увеличенная потребность в кислороде, ведущая к повышению уровня эритропоэтина, наблюдается при различных формах анемий. Однако продукция эритропоэтина и ответ на него эритропоэза неоднозначны при разных формах анемии и зависят от множества факторов. Например, значительное повышение эритропоэтина при апластической анемии в сыворотке и моче больных, возможно, обусловлено не только потребностью в нем, но и его пониженным потреблением. Вместе с тем потребность в кислороде может быть и сниженной. Например, белковое голодание приводит к снижению метаболизма и потребности в кислороде и в связи с этим – к уменьшению продукции эритропоэтина и эритропоэза, что проявляется в первую очередь в резком уменьшении ретикулоцитов в крови. Другим состоянием со снижением эритропоэза вследствие уменьшения потребности в кислороде и снижения продукции эритропоэтина является длительная гиподинамия (например, постельный режим, особенно с опущенной головой). Данное изменение эритропоэза можно наблюдать при эритремии. Регуляция миелопоэза Развитие и широкое распространение метода культивирования костного мозга и крови в агаровой культуре позволили более детально изучить регуляцию растущей в этой культуре бипотенциальной колониеобразующей гранулоцитарно-моноцитарной клетки-предшественницы (КОЕ-ГМ). Для роста колоний этой клетки-предшественницы в культуре и ее дифференцировки нужны особый колониестимулирующий фактор – КСФ или колониестимулирующая активность – КСА. Только лейкозные гранулоцитарно-моноцитарные клетки-предшественницы, в частности клетки миелоидного лейкоза мыши, могут расти без этого фактора. КСФ вырабатывается у человека моноцитарно-макрофагальными клетками крови и костного мозга, клетками плаценты, лимфоцитами, стимулированными определенными факторами, эндостальными клетками. КСФ представляет собой гликопротеин, он неоднороден по своему составу. Этот фактор состоит из двух частей: ЕО-КСФ (стимулирующего продукцию эозинофилов) и ГМ-КСФ (необходимого для продукции нейтрофилов и моноцитов). От концентрации КСФ зависит, продуцируются ли под его влиянием из одной клетки КОЕ-ГМ нейтрофилы или моноциты: для нейтрофилов необходима высокая концентрация КСФ, для моноцитов достаточно низкой концентрации. Продукция КСФ зависит от стимулирующих или ингибирующих влияний клеток, моноцитарно-макрофагальной и лимфоцитарной природы. Моноцитарно-макрофагальные элементы продуцируют вещества, подавляющие активность КСФ. К таким веществам-ингибиторам относятся лактоферрин, содержащийся в мембране макрофагов, и кислый изоферритин. Макрофаги синтезируют простагландины Е, которые прямо ингибируют (подавляют) КОЕ-ГМ. Т-лимфоциты также неоднородны в своем действии на КСФ и на КОЕ-ГМ. При истощении всех фракций Т-лимфоцитов в костном мозге и крови продукция КОЕ-ГМ повышается. При добавлении к такому костному мозгу лимфоцитов (но не Т-супрессоров) пролиферация КОЕ-ГМ повышается. Т-супрессоры костного мозга подавляют пролиферацию КОЕ-ГМ. Таким образом, в норме продукция КСФ, КОЕ-ГМ и ее потомства регулируется по системе обратной связи: одни и те же клетки являются и стимуляторами, и ингибиторами своей продукции. Основная масса клеток-предшественниц (которые составляют ничтожный процент от общего количества миелокариоцитов) производится «на всякий случай» и погибает неиспользованной. Однако само по себе постепенное повышение чувствительности к поэтинам позволяет отвечать дозированным увеличением необходимой в данный момент продукции. Если кровопотеря невелика, то в кровь выбрасывается дополнительно немного эритропоэтина, концентрация которого достаточна лишь для стимуляции КОЕ-Э. При тяжелой аноксии выброс эритропоэтина будет увеличен, и его концентрации хватит для стимулирования уже и более ранних предшественников эритропоэза, что позволит увеличить конечную продукцию эритроцитов на 1–2 порядка. Сходная картина наблюдается в гранулопоэзе. Содержание нейтрофилов и моноцитов в крови регулируется в основном колониестимулирующим фактором, большое количество которого ведет к повышению продукции нейтрофилов, а малое – к моноцитозу. Накопление моноцитов, в свою очередь, способствуя выработке простагландинов, изоферритина, подавляет продукцию колониестимулирующего фактора, и уровень нейтрофилов в крови снижается. Функции клеток крови В организме кровь выполняет множество функций:
Кровь также регулирует поступление к тканям и органам питательных веществ и поддерживает постоянство внутренней среды. Транспортная функция заключается в переносе большинства биологически активных веществ с помощью белков плазмы (альбуминов и глобулинов). Дыхательная функция осуществляется в виде транспорта кислорода и углекислого газа. Питательная функция заключается в том, что кровь доставляет ко всем органам и тканям питательные вещества – белки, углеводы, липиды. За счет наличия высокой теплопроводности, высокой теплоотдачи и способности легко и быстро перемещаться из глубоких органов к поверхностным тканям кровь регулирует уровень теплообмена организма с окружающей средой. Через кровь к местам выделения доставляются продукты метаболизма. Органы кроветворения и кроверазрушения поддерживают на постоянном уровне различные показатели, т. е. обеспечивают гомеостаз. Защитная функция заключается в участии в реакциях неспецифической устойчивости организма (врожденный иммунитет) и приобретенном иммунитете, системе фибринолиза за счет наличия в составе лейкоцитов, эритроцитов и тромбоцитов. Глава 2. Костный мозг Закладка костного мозга у эмбриона человека завершается к концу 3-го месяца внутриутробного развития, хотя в этот период он еще не принимает участия в процессе кроветворения. После окончания закладки костного мозга со стороны фиброзного слоя соединительной ткани в хрящевую ткань начинает прорастать ткань зародыша, богатая кровеносными сосудами. Эндотелий (внутренняя оболочка) эмбриональных сосудов частично разрушает хрящевую ткань, приводя тем самым к формированию системы полостей, которые носят название костномозговых пространств. В дальнейшем происходит процесс созревания клеток по трем направлениям: ретикулярные клетки, остеобласты и клетки жировой ткани. К началу второго триместра внутриутробного развития в костном мозге уже присутствуют элементы лимфоидной ткани, а также родоначальные клетки крови. С 5-го месяца внутриутробной жизни процесс костномозгового кроветворения приобретает определенную направленность с присутствием элементов гранулоцитарного, эритроцитарного и мегакариоцитарного рядов. В первой половине внутриутробной жизни ребенка процесс образования клеток крови происходит не только в костном мозге, но также в печени и селезенке. Клеточный состав костного мозга Костный мозг содержит в себе 2 группы клеток, к которым относятся клетки стромы, составляющие меньшинство, а также клетки паренхимы костного мозга в совокупности со зрелыми клетками крови. Зрелые кровяные клетки берут свое начало от клеток паренхимы костного мозга. При этом первая группа клеток костного мозга составляет меньшую его часть. Ретикулярная строма костного мозга включает в себя следующие клетки: фибробласты, остеобласты, эндотелиальные клетки (образующие внутреннюю оболочку кровеносных сосудов) и клетки жировой ткани. При гистологическом исследовании ткани костного мозга можно увидеть, что фибробласты содержат в себе круглое или вытянутой формы ядро с плотной или разреженной структурой хроматина, а также отростчатую цитоплазму. При цитологическом исследовании этого же препарата невозможно с достоверностью выделить фибробласт среди других клеток, входящих в состав костного мозга. Однако фибробласты могут внешне быть неотличимы от элементов лимфоидной ткани. В культуре фибробласты отличаются от других клеток круглым ядром, имеющим правильную грубоватую структуру хроматина и одно ядрышко. Фибробласты, содержащие компактные ядра без нуклеол, носят название фиброцитов. При гистологическом исследовании ткани костного мозга жировые клетки определяются очень хорошо. Как правило, в них заметны эксцентрично расположенное ядро небольшого размера, а также большое количество бесцветной цитоплазмы с отсутствием в ней других органелл. Жировые клетки можно легко увидеть при предварительной окраске ткани костного мозга определенными красителями, например суданом. Эндотелиальные клетки также не представляется возможным увидеть при микроскопическом исследовании ткани костного мозга. При гистологическом исследовании препарата видно, что данные клетки образуют внутреннюю поверхность всех кровеносных сосудов. Внутри эндотелиоцитов располагаются вытянутой формы ядро, большое количество цитоплазмы, имеющей беззернистую структуру. В процессе костеобразования принимают участие такие клетки, как остеобласты. Их диаметр составляет в среднем 20–25 мкм. Как правило, остеокласты имеют удлиненную или неправильную форму, округлой или овальной формы ядро, которое располагается эксцентрично и содержит в себе маленькое ядрышко. Остеобласты образуют эндостальную поверхность костномозговых полостей, отграничивая тем самым костный мозг от кости. В результате таких механизмов остеобласт образует вокруг себя кость, оказывается внутри нее и трансформируется там в остеоцит. В большом проценте случаев для постановки правильного диагноза при патологии крови прибегают к такому исследованию, как миелограмма. Миелограмма (от греч. myelos– «костный мозг» + gramma – «запись») является итогом прижизненного изучения клеточного состава ткани костного мозга, взятой от больного методом пункции. Миелограмма отражает как качественный, так и количественный состав клеток костномозговой ткани, который выражается в форме таблицы или диаграммы. Ее получают при изучении ткани костного мозга под микроскопом и используют в качестве метода диагностики большого количества заболеваний гематологического профиля. При проведении такого исследования, как миелограмма, как правило, выделяют ретикулярные клетки, включающие в себя все клеточные элементы, не имеющие места в рядах кроветворения. В ретикулярные клетки включены следующие элементы: доноры железа (в случае гемолитической анемии их процентное содержание может быть достаточно высоким), трудно идентифицируемые стромальные клетки, иммунобласты (встречаются в составе костного мозга крайне редко) и ряд других неидентифицируемых клеточных элементов. В случае какой-либо патологии со стороны системы крови, прежде всего при острых лейкозах, в миелограмме можно обнаружить достаточно большое количество клеток, которые являются атипичными по своей структуре, в результате чего их не представляется возможным идентифицировать. Одновременно с этим именно такие атипичные клетки, входящие в состав ткани костного мозга, имеют первостепенное значение для постановки диагноза. В том случае, если содержание атипичных клеток в составе костного мозга превышает 2%, необходимым является их точное морфологическое описание. Главной особенностью таких клеток является однородная структура ядра и цитоплазмы. Структурные элементы паренхимы костного мозга Миелобласты являются родоначальниками нейтрофилов, эозинофилов и базофилов. В обычном костном мозге эозинофильные и базофильные миелобласты неразличимы. Базофильные миелобласты определяются в случае хронического миелолейкоза, когда в крови содержится большое количество зрелых базофилов. Эозинофильный миелобласт может встретиться при высоких реактивных эозинофилиях, хотя это наблюдается крайне редко. Строение ядер всех 3 типов миелобластов идентично. Размер миелобласта составляет примерно 15–20 мкм. Ядро данной клетки имеет округлую форму, нежно-сетчатую структуру хроматина с равномерной окраской и одинаковым калибром нитей. Нейтрофильный промиелоцит отличается от миелобласта более крупными размерами, достигая в диаметре 25 мкм, а иногда и больше. Ядро промиелоцита сохраняет в себе остатки нежной структуры, но не имеет равномерной окраски и калибра нитей хроматина. В нем можно различить мелкие ядрышки. Для данной клетки характерна некоторая вариабельность, хотя она выделяется в качестве самой крупной среди гранулоцитов. Нейтрофильные миелоциты разделяют на крупные материнские (незрелые) миелоциты, диаметр которых может достигать 14–16 мкм, и дочерние – зрелые, меньших размеров, возникающие из материнских. Миелоцит обладает способностью к делению и переходу в следующую по зрелости клетку – метамиелоцит. Начиная с метамиелоцита кровяные клетки лишаются способности к делению. За ними сохраняется только способность к дальнейшему созреванию. Ядро материнского миелоцита чаще имеет овальную форму, в то время когда у дочернего миелоцита оно может иметь бобовидную форму с бухтообразным вдавлением, иногда оно является круглым. Структура ядра находится в прямой зависимости от степени зрелости миелоцита. Ядрышки в миелоцитах неразличимы, их можно выявить только при проведении специальной окраски. Цитоплазма миелоцита – светло-розового или светло-фиолетового цвета. При подсчете миелограммы материнский и дочерний миелоциты суммируют, что связано с отсутствием особого значения при их разделении для нормы и патологии. Нейтрофильный метамиелоцит носит название юного нейтрофила. Данная клетка может достигать в диаметре 12–13 мкм. В случае патологии со стороны системы крови все они могут становиться значительно крупнее и достигать в диаметре 20–22 мкм. Цитоплазма метамиелоцита нежно-розового цвета, заполнена мелкой специфической зернистостью, которая окрашивается в коричневато-розовые оттенки. Палочкоядерный нейтрофил представляет собой следующую ступень развития клетки нейтрофильного ряда. Ядро данной клетки может иметь разнообразную форму, в большинстве случаев оно вытянуто в виде палочки либо изогнуто в виде подковы. В отличие от ядра метамиелоцита оно вдвое тоньше. Палочкоядерный нейтрофил трансформируется в сегментоядерный путем разделения ядра на несколько связанных между собой сегментов. Эозинофильный промиелоцит в обычном костном мозге далеко не всегда можно отличить от нейтрофильного. Принадлежность промиелоцита к эозинофильному можно определить только лишь по особенностям его зернистости, которая является крупной и равномерно заполняет цитоплазму. Эозинофильный миелоцит. Для ядра данной клетки костного мозга специфична зернистость желто-красноватого цвета, густо заполняющая цитоплазму. Некоторые зерна остаются недозревшими, за счет чего при окраске принимают коричневатый оттенок. Эозинофильный метамиелоцит. Определить рассматриваемую клетку можно без особых затруднений по характерным контурам ее ядра, а также по характерной зернистости. Трудности возникают в том случае, когда густая зернистость заграждает ядро, контуры которого стушевываются. Палочкоядерный эозинофил содержит в себе ядро, изогнутое в виде подковы либо в виде другой фигуры. Ядро сегментоядерного эозинофила, как правило, содержит всего лишь 2 сегмента. При высоких эозинофилиях число сегментов увеличивается. Базофильный промиелоцит можно определить только в том случае, если удается увидеть характерную для него зернистость. Базофильный миелоцит. Данная клетка аналогично другим клеткам гранулоцитарного ростка той же степени зрелости одной из первых приобретает специфическую зернистость. Крупная зернистость обычно негусто заполняет цитоплазму рассматриваемой клетки. По структурным особенностям ядра, свойственным для всех миелоцитов, можно довольно легко определить базофильный миелоцит. Ядра зрелых базофилов обладают немного расплывчатыми очертаниями, имеют 3– или 4-лопастное строение. Диаметр базофила в норме не превышает 12–13 мкм. Тучные тканевые клетки также могут присутствовать в костном мозге, хотя более часто их определяют в ткани лимфатических узлов и селезенки. Эти элементы обладают характерной зернистостью красно-фиолетового цвета, что отличает их от темно-синей зернистости базофилов. Помимо того, зернистость тучных клеток выделяется богатым и густым расположением в цитоплазме. В цитоплазме также располагается округлое или овальное ядро. Функция тучных клеток заключается в выработке гепарина (гепариноциты) и синтезе гистамина. Роль родоначальной клетки лимфатического ряда принадлежит лимфобласту, диаметр которого достигает 15 мкм и больше. Ядро лимфобласта округлое, реже – несколько овальное. В ядре, как правило, находится от 1 до 2 ядрышек. Цитоплазма содержит зону просветления. Лимфобласт можно определить достоверно только при проведении цитохимического исследования. Пролимфоцит имеет грубую рыхлую структуру ядра, меньшие размеры, относительно большое количество цитоплазмы, что позволяет с легкостью отличать эту клетку от лимфобласта. В ряде случаев в ядре четко видно ядрышко. Пролимфоцит, как и зрелый лимфоцит, может содержать в цитоплазме малочисленную зернистость. Лимфоцит. Его ядро при изучении в световой микроскоп имеет округлую форму. Иногда определяется бобовидное вдавление. Цитоплазма окружает ядро неравномерным слоем, а иногда вообще едва заметна. Диаметр лимфоцита обычно составляет 8–9 мкм, хотя может достигать 12–15 мкм. Плазматические клетки в ткани костного мозга, взятой из грудины методом пункции, определяются всегда. Они происходят из В-лимфоцитов через стадию плазмобласта. В костном мозге плазмобласт определяют в редких случаях. Его диаметр в среднем составляет 16–20 мкм. Ядро захватывает большую долю клетки. Проплазмоцит характеризуется эксцентрично размещенным ядром, в котором далеко не всегда можно выявить ядрышко. Окраска цитоплазмы может иметь сероватый оттенок. Плазмоцит представляет собой зрелую плазматическую клетку со специфическими особенностями. Ядро колесовидной структуры обычно размещено эксцентрично. Цитоплазма нередко ячеистая, допустим клазматоз – отделение частиц цитоплазмы в ее периферических отделах. Монобласт – первая клетка моноцитарного ряда. Он не всегда отличим от миело– и лимфобласта. Как и эти клетки, монобласт содержит в себе ядро с 2–4 ядрышками. Цитоплазма нежно-голубого цвета. Если ядро монобласта имеет бобовидную форму или неправильные волнистые очертания, то установить природу клетки довольно легко. Промоноцит. Через стадию промоноцита формируется зрелая клетка данного ряда. Его ядро выделяется более грубой структурой хроматина, ядрышек не заметно. В цитоплазме можно определить мелкую пылевидную зернистость. Моноцит. В процессе дифференцировки промоноцита в моноцит на ядре появляются бухтообразное вдавление, а также волнистые контуры цитоплазмы, видимые при помощи как светового, так и электронного микроскопа. Вдавление может принять самые замысловатые формы. Цитоплазма, богатая пылевидной зернистостью, имеет своеобразный оттенок. Диаметр клеток моноцитарного ряда может составлять от 12 до 20 мкм. Макрофаги отличаются от других клеток костного мозга большими размерами. Их ядро сравнительно небольшого размера, округлой или слегка овальной формы, имеет сетчато-петлистую структуру. В нем заметно одно, в более редких случаях – два ядрышка. Широкий обод цитоплазмы имеет неправильные черты, окрашивается в светло-голубые или серо-голубые тона. Макрофаги содержат включения в виде обломков клеток, эритроцитов, пигмента, капель жира, а иногда даже бактерий. Липофаги попадаются в случае ксантоматоза, диабета, липидемии. Их диаметр достигает 40 мкм, цитоплазма имеет ячеистое строение. При обыкновенной окраске с фиксацией в спирте ячеистость определяется в виде пустот округлой формы («соты») разного размера. Ядро липофага часто оттеснено и деформировано. Остеокласты относятся к классу макрофагов. Во взрослом организме их появление связано с процессами рассасывания костной ткани. Они выявляются в случае опухолевых процессов, в месте локализации переломов костей. Остеокласты представляют собой гигантские клетки, диаметр которых достигает 40–80 мкм, содержат большое число ядер. Количество ядер, так же как и размеры клетки, подвержено значительным колебаниям. По морфологическим качествам остеокласты трудно ошибочно принять за иные гигантские клетки. Они схожи с гигантскими клетками инородных тел, с которыми обладают генетическим родством. Ядра остеокласта группируются скоплениями либо могут располагаться в цитоплазме равномерно. Диаметр ядер составляет около 10–12 мкм. В ядрах можно определить одиночное небольшого размера ядрышко. Цитоплазма окрашивается в нежные тона, время от времени приобретает пылевидную зернистость. Эритробласт – первая морфологически идентифицируемая клетка эритроидного ростка. Его ядро округлой формы и содержит в себе от 2 до 4 ядрышек. По морфологии эритробласт можно легко распознать. Его диаметр составляет 16–20 мкм. В процессе созревания клеток рассматриваемого ряда, начиная от эритробласта, размеры ядра, так же как и клетки в целом, уменьшаются. Пронормоцит включает отчетливо ограниченное округлой формы ядро и цитоплазму с околоядерным просветлением. Немного более грубая структура ядра, меньшие по сравнению с эритробластом размеры и отсутствие ядрышек, как правило, дают возможность дифференцировать пронормоцит от эритробласта. В случае патологии крови такие отличия не имеют значения. Следующим клеточным элементом в эритроидном ряду являются нормоциты. В зависимости от степени насыщения гемоглобином они подразделяются на базофильные, полихроматофильные и оксифильные, или ортохромные, нормоциты. Гемоглобин скапливается в цитоплазме при непосредственном участии в этом процессе ядра. Этот факт подтверждается первоначальным появлением гемоглобина вокруг ядра. Непрерывное накопление гемоглобина в цитоплазме клетки обусловливает восприимчивость как кислых, так и основных красок. При абсолютном насыщении клетки гемоглобином цитоплазма при окрашивании приобретает розовый цвет. Такой тип клеток носит название оксифильного нормоцита. При многих типах анемий в кровь выходят незрелые эритроциты. До выхода из костного мозга эритроциты 2–4 дня задерживаются в нем. После циркуляции в крови около 2 суток они трансформируются в зрелые эритроциты. Весь цикл формирования от эритробласта до нормоцита и эритроцита составляет более 100 ч. Мегакариоциты представляют собой гигантские клетки костного мозга, размером 60–120 мкм. Ядро их полиморфно, образовано фрагментами различной формы. Большая цитоплазма обильна зернистостью, имеющей определенное значение для созревания тромбоцитов. В нормальном костном мозге можно заметить образование и отхождение тромбоцитов от цитоплазмы мегакариоцита. Мегакариоциты подразделяются на незернистые, зернистые и обильнозернистые. Для мегакариоцита роль материнской клетки выполняет мегакариобласт. Его ядро имеет 8–10 мкм в диаметре. В ядре мегакариоцита определяются ядрышки. Цитоплазма охватывает ядро незначительным пояском и не включает зернистости. Промегакариоцит по своим размерам существенно больше мегакариобласта. Размеры цитоплазмы могут выраженно превалировать над размером ядра. Ядро имеет неровные очертания, ядрышки отсутствуют. Выделение данной клетки является условным. Тромбоциты, или кровяные пластинки. Данные клетки крови не являются истинными клеточными образованиями. Они имеют четко организованные элементы (ядро и цитоплазму) только лишь у низших позвоночных. Диаметр тромбоцитов составляет 2–4 мкм. В их структуре имеется центральная зона – грануломер, который окрашивается в фиолетово-красные тона, а также периферическая часть – гиаломер. Тромбоциты делятся на юные, зрелые и старые. В отличие от юных тромбоцитов, обладающих расплывчатыми контурами, зрелые имеют округлую форму с четкими границами гиаломера и хорошо выраженным грануломером. Старые формы тромбоцитов имеют гораздо меньший размер. В диагностике многообразных заболеваний кроветворной системы немаловажную роль играет цитологическое исследование костного мозга, т. е. определение его клеточного состава. Прежде чем приступить к изучению костного мозга в микроскопе, необходимо предварительно просмотреть препарат на малом увеличении. Такой подход позволяет установить, насколько препарат костного мозга обеспечен клеточными элементами, а также обнаружить скопления клеточных элементов, похожих на опухолевые. С целью определения клеточного состава костного мозга в процентном отношении необходимо произвести подсчет не менее 400 клеток. Состав костного мозга может колебаться в довольно широких пределах. Для подсчета мегакариоцитов и определения клеточного состава костного мозга пунктат подсчитывают таким же образом, как считают лейкоциты. Лейкоэритробластическое соотношение устанавливает отношение элементов белого и красного ряда. При отсутствии патологии оно составляет 4 (3) : 1. В случае полной функциональной полноценности костного мозга увеличение клеток красного ряда тем выше, чем больше выражена анемия по показателям периферической крови. При оценке функционального состояния костного мозга немаловажное значение отводится его «барьерной» функции. Глава 3. Моноциты и макрофаги Моноциты и макрофаги являются основными клетками системы фагоцитирующих мононуклеаров (ВОЗ) или макрофагальной системы И. И. Мечникова. Моноциты берут начало от гранулоцитарно-моноцитарной клетки-предшественницы, макрофаги – от моноцитов, переходящих из кровяного русла в ткани. Макрофаги присутствуют во всевозможных тканях человеческого организма: в костном мозге, в соединительной ткани, в легких (альвеолярные макрофаги), в печени (купферовские клетки), в селезенке и лимфатических узлах, в серозных полостях (брюшной полости, полости плевры, полости перикарда), в костной ткани (остеокласты), в нервной ткани (микроглиальные клетки), в коже (клетки Лангерганса). Они могут быть как свободными, так и фиксированными. Кроме того, к макрофагальным элементам относятся и дендритические клетки (имеющие большое количество коротких ветвящихся отростков), присутствующие во всех тканях. При проведении многочисленных операций по трансплантации костного мозга от донора иного пола было доказано кроветворное происхождение альвеолярных макрофагов, купферовских клеток, клеток Лангерганса и остеокластов. Сформировавшись в костном мозге, моноцит находится там от 30 до 60 ч. После этого он делится и поступает в системный кровоток. Период циркуляции моноцита в крови составляет приблизительно 72 ч, где происходит его созревание. Ядро моноцита трансформируется из круглого сначала в бобовидное, а затем в лапчатое. Помимо этого, отмечается изменение структуры генетического материала клетки. Цвет цитоплазмы моноцита может быть совершенно различным – от базофильного до серо-голубого или даже розоватого. После выхода из кровяного русла моноцит больше не может вернуться в системную циркуляцию. Макрофаги, расположенные в различных тканях человеческого организма, имеют ряд общих особенностей. При исследовании альвеолярных макрофагов было выявлено, что тканевые макрофаги поддерживают свою популяцию не только за счет образования в костном мозге, но также за счет имеющейся у них способности к делению и самоподдержанию. Данная отличительная черта макрофагов становится очевидной в случае подавления образования данных клеток крови в костном мозге под влиянием облучения или препаратов с цитостатическим действием. Ядро макрофага имеет овальную форму. Цитоплазма клетки достаточно большая, не имеет четких границ. Диаметр макрофага в норме колеблется в широких пределах: от 15 до 80 мкм. Специфическими функциональными признаками макрофагов служат способность прилипать к стеклу, поглощение жидкости и более твердых частиц. Фагоцитоз – «пожирание» чужеродных частиц макрофагами и нейтрофилами. Данное свойство клеток организма открыл И. И. Мечников в 1883 г.; он же предложил указанный термин. Фагоцитоз складывается из захвата клеткой чужеродной частицы и заключения ее в пузырек – фагосому. Образовавшаяся структура продвигается вглубь клетки, где переваривается при помощи ферментов, высвобождающихся из особых органелл – лизосом. Фагоцитоз представляет собой наиболее древнюю и важную функцию макрофагов, благодаря которой они избавляют организм от чужеродных неорганических элементов, разрушенных старых клеток, бактерий, а также иммунных комплексов. Фагоцитоз – одна из основных систем защиты организма, одно из звеньев иммунитета. В макрофагах его ферменты, так же как многие другие структуры, подчинены роли данных кровяных клеток в иммунитете и в первую очередь – фагоцитарной функции. В настоящее время известно более 40 веществ, продуцируемых микрофагом. Ферментами моноцитов и макрофагов, реализующими переваривание образующихся фагосом, являются пероксидаза и кислая фосфатаза. Пероксидаза содержится только в таких клетках, как монобласты, промоноциты и незрелые моноциты. В клетках последних двух стадий дифференцировки пероксидаза присутствует в очень малом количестве. Зрелые клетки и макрофаги настоящий фермент, как правило, не содержат. Содержание кислой фосфатазы увеличивается в процессе созревания моноцитов. Наибольшее ее количество – в зрелых макрофагах. Из поверхностных маркеров моноцитов и макрофагов иммунному фагоцитозу способствуют рецепторы к Fc-фрагменту иммуноглобулина G и к компоненту комплемента С3. С помощью указанных маркеров на поверхности моноцитарно-макрофагальных клеток закрепляются иммунные комплексы, антитела, различные клетки крови, покрытые антителами или комплексами, состоящими из антитела и комплемента, которые затем втягиваются внутрь клетки, осуществляющей фагоцитоз, и перевариваются ею либо сохраняются в фагосомах. Кроме фагоцитоза, моноциты и макрофаги обладают способностью к хемотаксису, т. е. способны двигаться в направлении разности содержания определенных веществ в клетках и вне клеток. Также данные кровяные клетки могут переваривать микробы и продуцировать несколько компонентов комплемента, играющих ведущую роль в образовании иммунных комплексов и в активации лизиса антигена, продуцировать интерферон, ингибирующий размножение вирусов, секретировать особый белок лизоцим, обладающий бактерицидным действием. Моноциты и макрофаги продуцируют и секретируют фибронектин. Данное вещество является по своей химической структуре гликопротеидом, связывающим продукты клеточного распада в крови, играющим важную роль во взаимодействии макрофага с иными клетками, в прикреплении (адгезии) на поверхности макрофага элементов, подлежащих фагоцитозу, что связано с наличием на мембране макрофага рецепторов к фибронектину. С защитной функцией макрофага связана также его способность продуцировать эндогенный пироген, представляющий собой специфический белок, который синтезируется макрофагами и нейтрофилами в ответ на фагоцитоз. Выделяясь из клетки, данный белок оказывает влияние на центр терморегуляции, расположенный в головном мозге. В результате повышается установленная указанным центром температура тела. Обусловленное воздействием эндогенного пирогена повышение температуры тела способствует борьбе организма с инфекционным агентом. Способность к выработке эндогенного пирогена увеличивается по мере созревания макрофагов. Макрофаг не только организует систему неспецифического иммунитета, заключающуюся в защите организма от любого инородного вещества или клетки, постороннего для данного организма или ткани, но и принимает непосредственное участие в специфическом иммунном ответе, в «представлении» чужеродных антигенов. Данная функция макрофагов связана с существованием на их поверхности особого антигена. HLA-DR-белок играет предопределяющую роль в развитии специфического иммунного ответа. У человека существует 6 вариантов молекулы HLA-DR-подобного белка. Этот белок присутствует практически у всех кроветворных клеток, начиная от уровня полипотентных клеток-предшественниц, но отсутствует на зрелых элементах, имеющих кроветворную природу. HLA-DR-подобный белок определяется и у эндотелиальных клеток, и у сперматозоидов, и у многих других клеток человеческого организма. На поверхности незрелых макрофагов, имеющихся преимущественно в тимусе и селезенке, также присутствует HLA-DR-подобный белок. Самое большое содержание такого белка обнаружено на дендритических клетках и на клетках Лангерганса. Такие макрофагальные клетки являются активными участниками иммунного ответа. Чужеродный антиген, попадающий в организм человека, адсорбируется поверхностью макрофага, поглощается им, оказываясь на внутренней поверхности мембраны. Затем антиген расщепляется в лизосомах. Фрагменты расщепленного антигена выходят из клетки. Часть этих фрагментов антигена взаимодействует с молекулой HLA-DR-подобного белка, в результате чего образуется комплекс на поверхности макрофага. Такой комплекс выделяет интерлейкин I, поступающий к лимфоцитам. Этот сигнал воспринимается Т-лимфоцитами. У Т-лимфоцита-амплифайера возникает рецептор к HLA-DR-подобному белку, ассоциированному с фрагментом чужеродного антигена. Активированный Т-лимфоцит выделяет второе сигнальное вещество – интерлейкин II и ростовой фактор для лимфоцитов всех типов. Интерлейкин II активирует Т-лимфоциты-хелперы. Два клона лимфоцитов данного типа отвечают на действие чужеродного антигена, продуцируя фактор роста В-лимфоцитов и фактор дифференцировки В-лимфоцитов. Результатом активации В-лимфоцитов является продукция специфических к данному антигену иммуноглобулинов-антител. Таким образом, несмотря на то что распознавание чужеродного антигена является функцией лимфоцитов без участия макрофага, переваривающего антиген и соединяющего часть его с HLA-DR-подобным белком поверхности, невозможны представление антигена лимфоцитам и иммунный ответ на него. Макрофаги обладают способностью переваривать не только бактериальные клетки, эритроциты и тромбоциты, на которых фиксированы некоторые компоненты комплемента, в том числе стареющие или патологически измененные, но также и опухолевые клетки. Такой вид активности макрофагов получил название тумороцидной. Из этого нельзя сделать вывод о действительной борьбе макрофагов с опухолью, а именно «признании» ими такого типа клеток как чужеродной ткани, в связи с тем что в любой опухоли присутствует очень много стареющих клеток, подлежащих фагоцитозу аналогично всем неопухолевым стареющим клеткам. Отдельные факторы, продуцируемые клетками моноцитарно-макрофагальной природы (например, простагландины Е, лизоцим, интерферон), участвуют и в иммунной функции, и в кроветворении. Кроме того, макрофаги помогают развитию эозинофильной реакции. Доказана макрофагальная природа остеокластов. Макрофаги способны, во-первых, непосредственно растворять костную ткань, во-вторых, стимулировать продукцию остеокластстимулирующего фактора Т-лимфоцитов. Данная функция макрофагов может оказаться ведущей в патологии, обусловленной опухолевой и реактивной пролиферацией макрофагов. Весьма существенную роль играют макрофаги в постоянстве внутренней среды. Прежде всего они являются единственными клетками, продуцирующими тканевой тромбопластин, и запускают сложный каскад реакций, обеспечивающих свертывание крови. Однако, по-видимому, повышение тромбогенной активности в связи с жизнедеятельностью макрофагов может быть обусловлено также обилием как секретируемых ими, так и внутриклеточными, выделяемыми при распаде клеток, протеолитических ферментов, продукцией простагландинов. Вместе с тем макрофаги продуцируют активатор плазминогена – антисвертывающий фактор. Глава 4. Эозинофилы Эозинофилы представляют собой особый класс гранулоцитов, отличающихся своим происхождением, строением, спектром ферментов, кинетикой и своеобразной ролью в адаптационных реакциях. Строение Зрелый эозинофил имеет диаметр 12–17 мкм, двухлопастное ядро и гранулы оранжево-красного цвета. При созревании эозинофил проходит те же стадии, что и нейтрофил. В цитоплазме эозинофилов по мере созревания появляются гранулы 2 типов. Большие овальной формы эозинофильные гранулы образуются на ранних стадиях развития, а на более поздних стадиях приобретают кристаллическую структуру и окрашиваются анилиновыми красителями. Малые гранулы эозинофилов, однородные по структуре, появляются на стадии метамиелоцита. Мембрана эозинофила несет специфические антигенные структуры, определяемые с помощью специальных сывороток. На поверхности эозинофилов обнаружены рецепторы для особых белков – антител, обозначаемых как иммуноглобулины класса G – IgG. Количество таких иммуноглобулинов значительно увеличивается при активации клетки. Существование рецепторов для иммунных комплексов, содержащих белки-иммуноглобулины класса Е – IgE, было доказано с помощью стимуляции активности эозинофилов после взаимодействия их с иммунной сывороткой. Характерной особенностью мембраны эозинофилов являются рецепторы для С4 и С3 компонентов системы комплемента. Следует отметить, что на мембране нейтрофилов имеются рецепторы для С3 компонента комплемента. Количество рецепторов для комплемента на эозинофилах увеличивается в период их участия в иммунных реакциях, при этом параллельно возрастает их токсическое действие на патологические чужеродные клетки. Ферменты Пероксидаза эозинофилов отличается от пероксидазы нейтрофилов более низкой бактерицидной активностью. Арилсульфатаза, содержащаяся главным образом в мелких гранулах эозинофилов, инактивирует вещества, способствующие развитию аллергической реакции немедленного типа. Уровень этого фермента в эозинофилах в 15 раз выше, чем в нейтрофилах. Фосфолипаза D нейтрализует фактор активации тромбоцитов, уменьшает способность тромбоцитов к выделению веществ, образующихся в их гранулах. Количество фосфолипазы D в эозинофилах человека в 10 раз больше, чем в нейтрофилах. Эозинофилы содержат также намного большее количество других ферментов: липофосфорилазы, пероксидазы, глюкуронидазы и кислой глицерофосфатазы по сравнению с нейтрофилами. Важнейшими в функциональном отношении белковыми структурами эозинофилов являются большой основной (щелочной) белок и катионный белок эозинофилов. Большой основной белок составляет около половины белков больших гранул. Он обладает сродством к анилиновым красителям, определяющим окраску этих гранул, способен нейтрализовать гепарин, повреждать личинки ряда паразитов и некоторые клетки организма, т. е. обладает свойством цитотоксичности. В эксперименте показано участие катионных белков эозинофилов в воспалительных реакциях, влияние на плазменный гемостаз через калликреин-кининовую систему и XII фактор свертывания, повреждающее действие на эндотелий (внутреннюю оболочку кровеносных сосудов). Предполагают, что от этих белков зависит повреждение эндокарда (внутренней оболочки сердца) при длительных гиперэозинофилиях (чрезмерном увеличении количества эозинофилов). Происхождение и развитие эозинофилов Впервые в филогенезе эозинофилы обнаруживаются у высших позвоночных. У человека они появляются на 8-й неделе внутриутробного развития одновременно с формированием лимфоцитарной системы. Эозинофилы ведут свое происхождение от общей клетки – предшественницы миелопоэза. На ранних этапах дифференцировки эозинофильный росток обособляется от других ростков. Существует отдельная клетка – предшественница эозинофилопоэза, в развитии которой особое значение имеют эозинофилопоэтины (вещества, оказывающие непосредственное влияние на образование эозинофилов), вырабатываемые лимфоцитами селезенки. Иммунологический контроль четко прослеживается не только на первом, но и на всех остальных этапах развития эозинофилов. При культивировании костного мозга в присутствии лимфоцитов периферической крови в агаровых культурах эозинофилы образуют отдельные плотные колонии к 12-му дню культивирования. При отсутствии тимуса (вилочковой железы) у больных не наблюдаются эозинофильные реакции. Повышение уровня IgE, напротив, приводит к ускоренному созреванию эозинофильного ростка, который не касается других ростков миелопоэза. Кинетика эозинофилов Начальные стадии созревания эозинофилов в костном мозге длятся 34 ч, после этого клетки выходят в кровоток. В кровотоке эозинофилы находятся недолго, после чего располагаются главным образом в покровных тканях (коже, слизистых оболочках желудочно-кишечного тракта, дыхательных и мочеполовых путей). При острых воспалительных процессах значительное количество эозинофилов выходит из циркуляции и накапливается по периферии очага воспаления. Содержание эозинофилов в тканях человека приблизительно в 100 раз превышает таковое в кровотоке. При эозинофилиях кинетика эозинофилов значительно изменяется. На ранних стадиях созревания укорачиваются клеточные циклы, увеличивается митотический индекс, время генерации сокращается в 3 раза, эозинофилы появляются в периферической крови в 2 раза быстрее. Из циркуляции эозинофилы исчезают в течение 3 ч, а затем вновь возвращаются в кровоток из тканей. Длительность рециркуляции является одной из причин эозинофилии (повышения содержания эозинофилов в крови). Миграция и активация эозинофилов происходят под влиянием хемотаксических факторов различного происхождения. Они представляют собой химические вещества, притягивающие к себе эозинофилы. При появлении данных веществ в кровотоке эозинофилы начинают двигаться в их направлении. Если имеется такая необходимость, то эозинофилы могут передвигаться против тока крови, т. е. ретроградно. Функциональные особенности эозинофилов Роль эозинофилов заключается в предупреждении проникновения антигена в сосудистое русло, а именно в генерализации иммунного ответа. Область реакции отграничивают эозинофилы с помощью нейтрализации продуктов обмена веществ (метаболитов), участвующих в уничтожении антигена. При образовании большого количества метаболитов место реакции отграничивается с помощью местного некроза и фиброзирования, что также является функцией эозинофилов. Таким образом, эозинофилы завершают иммунный ответ на уровне подслизистого и подэпителиального слоя, защищая организм от множества нецелесообразных общих иммунных реакций на небольшие дозы проникающих чужеродных антигенов. Этот процесс эозинофилы осуществляют вместе с IgE-антителами, базофилами, тучными клетками, макрофагами, лимфоцитами и комплементом. При патологических состояниях весьма своеобразно проявляются специфические функции эозинофилов, заключающиеся в нейтрализации метаболитов и активации процессов фиброзирования (образования фиброзной ткани в месте дефекта). Функциональная нагрузка у эозинофилов особенно велика при развитии аллергических реакций. Под влиянием хемотаксических факторов, выделяемых Т-лимфоцитами с повышенной чувствительностью, эозинофилы мигрируют к месту иммунной реакции на самых первых ее этапах. В период миграции и инфильтрации (пропитывания тканей) происходят усиленное созревание эозинофилов, накопление в них ферментов и увеличение количества рецепторов для С3, а также IgE-содержащих комплексов. Реализация аллергической реакции, заключающаяся во взаимодействии IgE-антител с аллергеном на поверхности тучных клеток и приводящая к выходу субстанций анафилаксии и гистамина, вызывает ответ эозинофилов. Данный ответ состоит в выделении серии инактивирующих ферментов, которые нейтрализуют гистамин, медленно реагирующие субстанции анафилаксии (состояние повышенной чувствительности организма к повторному введению того же аллергена), литический фактор тромбоцитов, гепарин, гистамин. Эозинофилы могут «пожирать» гранулы, выделяемые тучными клетками, препятствуя выходу из гранул гепарина и протеаз (ферментов). После всестороннего подавления аллергической реакции эозинофилы способствуют восстановлению тканевых тучных клеток, выполняя при этом восстановительную функцию. Таким образом, из всего вышесказанного можно сделать вывод, что в эозинофилах синтезируется огромное количество химических веществ, принимающих непосредственное участие в развитии аллергической реакции. Причем все эти вещества активно выделяются из гранул эозинофила при попадании в организм аллергена любого происхождения, препятствуя развитию аллергичекой реакции. Основная функция эозинофилов в противогельминтном (противоглистном) иммунитете состоит в цитотоксическом эффекте. Эта функция придает эозинофилам особое значение в защите организма. Активированный эозинофил, участвующий в реакции, имеет большое количество рецепторов для специфических IgE-антител. При контакте такого эозинофила с личинкой происходят его дегрануляция (выделение химических веществ из гранул) и отложение большого основного белка и пероксидаз (ферментов) на поверхности личинки. Цитотоксичность активированных эозинофилов по отношению к личинкам достигает 70–80%, в то время как у моноцитов и макрофагов она в 2–3 раза ниже. Гиперэозинофилия, наблюдаемая при миграции личинок, является основой противопаразитарной защиты при снижении функции Т-клеточного иммунитета и стимуляции синтеза IgE. Глава 5. Лимфоциты Лимфоциты представляют собой уникальную по разнообразию популяцию клеток, происходящих из различных предшественников и объединяемых единой морфологией. В световом микроскопе лимфоциты имеют форму мононуклеара, содержат одно ядро округлой формы и узкий ободок цитоплазмы. Морфологическими стадиями развития лимфоцита являются лимфобласт, иммунобласт, пролимфоцит и лимфоцит. У части лимфоцитов (В-лимфоцитов) конечными стадиями становятся плазмобласт, проплазмоцит и плазматическая клетка. Набор ферментов в лимфоцитах меняется в зависимости от принадлежности к определенной субпопуляции и функциональной активности. Подразделение лимфоцитов связано с их происхождением, функциональными особенностями и иммуноморфологической характеристикой. По происхождению лимфоциты подразделяются на две основные субпопуляции: Т-лимфоциты (тимусзависимые), предшественником которых является колониеобразующая клетка костного мозга, ее дифференцировка происходит под влиянием тимозина (гормона тимуса); В-лимфоциты, происходящие из колониеобразующей клетки костного мозга и развивающиеся под влиянием активаторов, не связанных с тимусом. Среди лимфоцитов периферической крови выделяется третья сборная группа, не имеющая основных признаков (маркеров) Т– и В-лимфоцитов и обозначаемая как «ни Т-, ни В-». Клетки, входящие в ее состав, по морфологической структуре идентичны лимфоцитам, но различаются по происхождению и функциональным особенностям. Функциональное подразделение лимфоцитов связано с их участием в иммунологической реакции:
Иммуноморфологическое подразделение лимфоцитов позволяет разграничить их по функциональной принадлежности и происхождению с помощью определения на мембране набора рецепторов и антигенов, различного у каждой субпопуляции. С помощью мембранных структур клетка «узнает» антиген и взаимодействует с другими иммунокомпетентными клетками. Комплекс антигенных и рецепторных структур мембраны лимфоцита является иммуноморфологической характеристикой клетки. В него входят иммуноглобулины, антигены гисто-совместимости, рецепторы для компонентов комплемента, гетерогенных эритроцитов, митогенов. Т-лимфоциты Т-лимфоциты представляют собой сложную систему различных в функциональном отношении клеток, объединяемых происхождением и присутствием на поверхности общего антигена – тимусного человеческого лимфоцитарного антигена. Предшественники Т-лимфоцитов, попадая в корковый слой тимуса, быстро размножаются, превращаясь в тимоциты. При дифференцировке тимоцитов образуются ранние, или менее зрелые, Т-лимфоциты и более зрелые Т2-лимфоциты. Идентифицировать ранние стадии дифференцировки можно только специализированными методами, а именно иммуноморфологически и гистохимически. Все ранние стадии Т-лимфоцита (до выхода из тимуса) содержат фермент – терминальную дезоксинуклеотидилтрансферазу, которая исчезает на уровне Т-лимфоцитов. Реакция на кислую фосфатазу на ранних стадиях определяется как интенсивная; фермент расположен в аппарате Гольджи. Кроме способности дифференцироваться в зрелые Т-лимфоциты, тимоциты несут определенную функциональную нагрузку в иммунном ответе. В присутствии тимоцитов иммунный ответ зрелых Т-лимфоцитов лимфатических узлов оказывается значительно более интенсивным. Количество вырабатываемых в тимусе Т-лимфоцитов очень велико – число мигрирующих за сутки из тимуса Т-клеток в 5–20 раз превышает их общее количество в периферической крови. Среди зрелых Т-лимфоцитов, образующихся после контакта с антигеном, различаются антигенреактивные клетки, хелперы, киллеры, эффекторы аллергической реакции, супрессоры, клетки иммунологической памяти, а также выделяется особый вид регулирующих Т-клеток, объектом действия которых являются стволовая клетка костного мозга и первые этапы ее дифференцировки. Большинство зрелых Т-лимфоцитов содержат в цитоплазме различные ферменты, включая нейтральную неспецифическую эстеразу, кислую фосфатазу и ?-глюкуронидазу. Т-лимфоциты отличаются от В-лимфоцитов высокой скоростью миграции, хотя постоянно рециркулирует только часть из них. Другие Т-клетки – «оседлые» и занимают свое место в периферических лимфатических органах. Кроме общих характеристик, свойственных всем зрелым Т-лимфоцитам, каждая субпопуляция Т-клеток имеет свои иммуноморфологические и функциональные особенности. Антигенреактивные Т-лимфоциты относятся к Т2-лимфоцитам. Они первыми реагируют на присутствие антигена, запускают в реакцию хелперы и супрессоры и способствуют их делению (пролиферации), но эффекторами не являются. Предполагают, что антигенреактивные Т-клетки, впервые реагирующие с антигеном, – предшественники клеток-амплификаторов. Антигенреактивные Т-клетки вместе с более ранними стадиями созревания Т-лимфоцитов составляют основную массу Т-лимфоцитов периферической крови и лимфы, им свойственна высокая способность к миграции (перемещению). После встречи с антигеном эта клетка превращается в иммунобласт, который, выделяя медиаторы, содействует запуску иммунной реакции в ближайшем лимфатическом узле. При отсутствии или резком уменьшении количества антигенреактивных Т-клеток нарушается процесс распознавания, что проявляется в снижении ответа на бактериальные, вирусные и грибковые антигены, появлении аутоиммунных расстройств. Этому способствуют отсутствие вилочковой железы, хроническая потеря лимфы из грудного протока, глубокая кахексия (истощение). Нарушение антигенного распознавания обнаружено при синдроме иммунной амнезии. При этом заболевании ослаблен клеточный и гуморальный иммунитет (приживление трансплантатов, отсутствие ответа на вакцинацию, сывороточные иммуноглобулины не обладают функцией антител, отсутствует IgА, нарушается соотношение лимфоцитов). Распознавание может быть нарушено изолированно в отношении белковых или полисахаридных антигенов или их сочетания. К антигенреактивным Т-клеткам относится и группа клеток иммунологической памяти. В функциональном отношении они отличаются тем, что узнают антиген в фазу вторичного иммунного ответа, при повторном контакте с антигеном, причем реагируют на антиген раньше и значительно интенсивнее, чем при первом контакте. Т-клетки иммунологической памяти относятся к потомкам хелперов, супрессоров или киллеров и несут тот же фенотип мембранных антигенов. Т-хелперы неоднородны. Более зрелые из них являются хелперами Т-В, функция которых заключается в воздействии на определенный клон В-лимфоцитов. Хелперы Т-Т, более ранние по дифференцировке, способствуют пролиферации Т-киллеров и эффекторов аллергической реакции. Воздействие Т-хелперов, преимущественно расположенных в селезенке и лимфатических узлах, на другие клетки осуществляется не только при непосредственном контакте, но и с помощью гуморальных медиаторов при обязательном участии макрофагов. Для выполнения основной задачи – представления антигена В-лимфоцитам в специальной связанной форме – рецепторы хелперов Т-В соединяются с антигеном, образуя комплекс, названный иммуноглобулином Т (IgT). В комплекс входят антиген и мономерная иммуноглобулиновая молекула, по биохимическим свойствам приближающаяся к классу IgM. Специфический хелперный фактор фиксируется на поверхности макрофага, способствуя концентрации молекул антигена. Для выработки фактора необходима большая доза антигена (чужеродного вещества). Т-хелперы вырабатывают хелперный фактор клеточного иммунитета. Его функция заключается в усилении цитотоксического действия и дифференцировки (созревания) киллеров, увеличении противоопухолевой активности макрофагов. Для стимуляции В-клеток необходимо совместное действие специфического и неспецифического хелперных факторов. Митогенный фактор Т-лимфоцитов выделяется вскоре после взаимодействия Т-хелперов с антигеном; он резко усиливает синтез ДНК в клетках, действуя неспецифически. Основные функции – поддержание пролиферации (деления) иммунокомпетентных клеток, усиление превращения лимфоцитов в иммунобласты (бласттрансформация), стимуляция выработки других медиаторов. Т-хелперы играют исключительно важную роль, определяя направление и силу иммунного ответа. Отмечены снижение количества хелперов и угнетение их функции при старении и опухолях. Содержание хелперов увеличено при аутоиммунных заболеваниях, системной красной волчанке, рассеянном склерозе, отторжении трансплантата. Хелперная функция Т-клеток зависит от их пролиферации и дифференцировки, поэтому легко подавляется ингибиторами белкового синтеза, в том числе цитостатиками. Т-эффекторы гиперчувствительности замедленного типа (ГЗТ) – название субпопуляции Т-клеток, в основном предназначенных для секреции лимфокинов. По происхождению Т-эффекторы ГЗТ относятся к той же группе предшественников, что и Т-хелперы, но способа их разделения пока не существует. Следует отметить, что гуморальные медиаторы синтезируются не только этими клетками, а практически всеми лимфатическими клетками и органами. К лимфокинам, источником которых в настоящее время считаются Т-эффекторы ГЗТ, относятся:
Лимфотоксин – фактор, образующийся под влиянием специфического антигена или митогена и выделяющийся без синтеза ДНК из активированной клетки. Лимфотоксин оказывает разрушающее действие на клетку через рецепторы мембраны, нарушая регуляторные механизмы и активируя внутриклеточные протеолитические системы. Основной объект действия лимфотоксина – измененная под влиянием вируса клетка. Иммунный интерферон синтезируется Т-клетками в присутствии Т-зависимых антигенов и митогенов. Его действие заключается в подавлении размножения вирусов, усилении ответа на Т-митогены, усилении фагоцитоза макрофагов, подавлении размножения и дифференцировки предшественников гранулоцитов, макрофагов, антителопродуцентов. Кроме перечисленных выше, Т-лимфоциты выделяют также остеокластактивирующий, плазминогенактивирующий факторы, а также простагландины. Совокупность гуморальных факторов, вырабатываемых как эффекторами ГЗТ, так и другими лимфоцитами, составляет вторую гуморальную иммунную систему организма. Основная задача лимфокинов состоит в обеспечении взаимодействия различных типов клеток и вовлечении их в иммунную реакцию. Лимфокины могут заменять функцию тех или иных субпопуляций лимфоцитов, обеспечивая при переносе реципиенту пассивный клеточный иммунитет, так же как и представители первой гуморальной иммунной системы (антитела). К настоящему времени в практике уже широко применяется лейкоцитарный интерферон, который, кроме противовирусного действия, может давать некоторый цитостатический эффект. Последний связан с влиянием интерферона на синтез ДНК и проявляется только при употреблении больших (токсических) доз препарата. Большинство Т-эффекторов ГЗТ находится в селезенке, хотя в некотором количестве они присутствуют и в других периферических лимфатических органах. Можно предположить, что в циркуляции есть главным образом предшественники Т-клеток иммунологической памяти ГЗТ, а зрелые продуценты гуморальных медиаторов локализуются в лимфатических органах. Т-супрессоры являются регуляторами направления и объема иммунной реакции, главным образом ограничивая деление клонов лимфатических клеток, угнетая антителообразование, дифференцировку киллеров, реакцию лимфоцитов в смешанной культуре, аллергический процесс и развитие ГЗТ. Под влиянием супрессоров развивается состояние иммунологической толерантности (иммуноареактивности) к антигену. Т-супрессоры неоднородны и различаются по происхождению и основной мишени воздействия. Супрессоры Т-клеток, как предполагают, происходят из более ранних Т-лимфоцитов, отличаются большой радиочувствительностью, действуют на предшественники Т-хелперов, причем осуществляют свою функцию без заметной пролиферации. Количество Т-супрессоров увеличивается с возрастом, особенно у женщин, изменяя соотношение между хелперами и супрессорами. Увеличивается количество Т-супрессоров, экспрессирующих антиген при инфекционном мононуклеозе и остром гепатите, а также при приживлении трансплантата. Большое значение Т-супрессоры имеют в связи с ростом опухоли. После инъекции супрессоров в эксперименте происходят подавление хелперов, киллеров, активированных макрофагов и усиление роста опухоли. При этом увеличивается селезенка, которая рассматривается как результат подавления всей популяции иммунокомпетентных клеток, в настоящее время может трактоваться как признак выраженной иммуносупрессии, сопровождающейся пролиферацией Т-супрессоров в селезенке. Увеличено количество супрессоров и при ряде врожденных иммунодефицитных состояний. Т-киллеры (цитотоксические Т-лимфоциты) являются основными эффекторными клетками, оказывающими цитотоксическое действие на клетки-мишени. Т-киллеры образуются в результате дифференцировки Т2-лимфоцитов после стимуляции клеточными антигенами. Основными антигенами, вызывающими клеточный цитотоксический ответ, служат HLA-ABC чужеродных или измененных клеток своего организма. Благодаря такой способности Т-киллеры уничтожают клетки трансплантата и мутантные клетки организма, в том числе опухолевые. В процессе развития иммунного ответа цитотоксическая реакция Т-клеток – одна из самых ранних и высокоспецифичных. Больше всего Т-киллеров в лимфатических узлах. Механизм действия Т-киллеров полностью не расшифрован. Известно, что при их контакте с клетками-мишенями не синтезируется ДНК, не происходит видимых изменений в клетке даже после их контакта с несколькими клетками-мишенями. Однако кратковременного контакта чужеродной клетки с Т-киллерами достаточно, чтобы вызвать необратимые изменения, вероятно, в результате осмотических нарушений в клетке-мишени. Цитотоксичность Т-киллеров может увеличиваться под влиянием различных гуморальных факторов и митогенов. Т-дифференцирующими (Td) названы лимфоциты, непосредственно влияющие на стволовые и колониеобразующие гемопоэтические клетки. В отдельный класс клеток, имеющих свой фенотип, Td еще не выделены. Не исключено, что Td представляют собой отдельный клон, включающий Т-хелперы, Т-супрессоры и другие типы лимфоцитов с определенной специфичностью к антигенам стволовых клеток. В любом случае особая роль в процессе кроветворения заставляет выделить Td в отдельную субпопуляцию. Как показали исследования в аллогенной системе, Td существенно влияют на дифференцировку гранулоцитарного и моноцитарного рядов. Снижение хелперного влияния на колониеобразование сказывается на соотношении количества эритроцитарных и гранулоцитарных колоний в сторону увеличения эритроцитарных. Дефицит Td-хелперов и повышение уровня Td-супрессоров имеют важное значение при старении организма, ряде врожденных иммунодефицитных состояний, развитии гипо– и апластических синдромов. Врожденные иммунодефицитные состояния, связанные с нарушением Т-клеточной системы иммунитета, нередко демонстрируют нарушения в системе кроветворения. При сочетании врожденного иммунодефицита с опухолью вилочковой железы отсутствует клеточный и гуморальный иммунитет, наблюдаются недоразвитие лимфатических узлов, уменьшение содержания лимфоцитов и плазмоцитов, а также недоразвитие лейкоцитарного и эритроцитарного ростков. Влияние Td-супрессоров у больных с апластической анемией выявлено при исследовании колониеобразования в культуре. Обработка Т-клеток больных анти-Т-сывороткой способствовала увеличению колониеобразования. Дифференцировка Т-лимфоцитов Пока не накоплено достаточно фактов, чтобы установить пути дифференцировки всех Т-клеток. С помощью моноклоновых антител достоверно выявлено направление созревания ранних стадий Т-лимфоцитов (тимоцитов, Т, и Т2). Пути антигензависимой дифференцировки зрелых Т-клеток, очевидно, неоднозначны. Например, Т-хелперы могут образовываться из Т– и Т2-лимфоцитов. То же относится и к Т-супрессорам. В-лимфоциты В-лимфоциты представляют собой систему клеток, объединяемых происхождением из костномозгового предшественника В-лимфоцитов. В функциональном отношении В-клетки, как и Т-лимфоциты, очень разнообразны. Среди В-клеток различают антителопродуценты (синтезирующие иммуноглобулины), киллеры, супрессоры, клетки иммунологической памяти. Все В-лимфоциты несут В-антиген. В-лимфоциты, как и Т-лимфоциты, проявляют способность к иммунному ответу на различных стадиях дифференцировки, а не только на уровне зрелых плазмоцитов. Первые этапы дифференцировки В-лимфоцита происходят в структурах костного мозга и являются антигеннезависимыми. Различают несколько этапов дифференцировки от стволовой клетки и общего предшественника лимфоцитов до зрелых В-лимфоцитов, выходящих из костного мозга и мигрирующих в периферические лимфатические органы для антителозависимого созревания. Самой первой стадией считают пре-В-лимфоцит, не имеющий цитоплазматических и поверхностных иммуноглобулинов, но обладающий В-антигеном и общим антигеном, свойственным острому лимфобластному лейкозу. Следующая стадия – пре-В-лимфоцит. На следующей стадии – ранних В-лимфоцитов – появляются молекулы Ig на мембране клетки, которые принадлежат к классу М. На стадии ранних В-лимфоцитов перестает определяться TdT в цитоплазме. Промежуточная стадия костномозгового В-лимфоцита не имеет общего лейкозного антигена. Следующие стадии дифференцировки В-лимфоцит проходит вне костного мозга: стадия зрелого лимфоцита отличается более низкой концентрацией В-антигена и В2-антигена и способностью к смене класса синтезируемых молекул Ig; конечным этапом дифференцировки В-лимфоцитов является плазматическая клетка, лишенная всех В-антигенов и поверхностных Ig и обладающая цитоплазматическими Ig в большой концентрации. Следует отметить, что все указанные стадии дифференцировки в полном объеме относятся только к В-лимфоцитам – антителопродуцентам, так как этапы созревания В-супрессоров и В-киллеров достоверно не установлены. Антигензависимая дифференцировка В-лимфоцитов происходит в периферических лимфатических органах в зародышевых центрах. Образование этих центров начинается сразу после рождения, хотя способность к синтезу антител класса М существует уже в эмбриональном периоде и проявляется при ряде внутриутробных инфекций. Подготовка к началу синтеза антител в период новорожденности заключается не только в быстром накоплении В-лимфоцитов, но и в подготовке плацдарма для их дифференцировки. Таким плацдармом становится селезенка с наибольшей относительной массой в период новорожденности, а количество фолликулов в ней быстро нарастает в течение первого года жизни. В них оседают В-лимфоциты – преимущественно «оседлые» клетки, они мигрируют меньше и значительно медленнее, чем Т-лимфоциты. Среди В-лимфоцитов наиболее многочисленны В-лимфоциты – антителопродуценты. Иммуноглобулины К иммуноглобулинам относятся белки животного происхождения, которые могут обладать активностью антител, а также белки, сходные с ними по химической структуре и, следовательно, по антигенной специфичности. В эту группу включены также белки, не имеющие активности антител, – миеломные белки, белки Бенс-Джонса, встречающиеся субъединицы иммуноглобулинов. Все Ig на 5 основных классов, обозначаемых символами IgG, IgА, IgM, IgD и IgE. Позже в классах IgG, А, M и D были определены подклассы, различающиеся по структуре и функциональным особенностям. Переключение синтеза IgM и IgG и IgA происходит непосредственно в антителосинтезирующих клетках. Механизм синтеза двух разных Ig предполагает одновременное функционирование двух генов. Клетка одного клона может синтезировать 2 Ig, различающихся по классу, но обладающих одной специфичностью. Нормальный гуморальный ответ всегда реализуется многими В-клеточными клонами, он поликлоновый. Это объясняется не только сложной структурой антигенов, но и перекрестным реагированием разных клонов на один антиген. Кроме того, реакция Т-хелперов на антигенную стимуляцию приводит к выделению как антигенспецифических, так и антиген-неспецифических растворимых хелперных факторов. Последние вызывают активацию многих клонов В-лимфоцитов и поликлоновый синтез антител. Вероятно, таким образом «перекрывается» весь возможный спектр антигенов, который во много раз превышает разнообразие узкоспециализированной клональной системы лифоцитов. Криоглобулины (криоиммуноглобулины) – это Ig, осаждающиеся при температуре ниже 37°С. Для выявления криоглобулинов сыворотку крови выдерживают в холодильнике при температуре 4°С в течение 72 ч. Для контроля за помутнением, гелификацией или выпадением осадка сыворотку просматривают каждые 24 ч. В ряде случаев криоглобулины выпадают в осадок при комнатной температуре или даже при температуре, близкой к температуре тела. Это может затруднять взятие крови из вены (следует пользоваться подогретой иглой) и исследование сыворотки (кровь гелифицируется в пробирке). Присутствие криоглобулинов контролируют, помещая сыворотку больного в термостат или на водяную баню при 37°С (осадок растворяется). Для изучения состава криоглобулинов осадок отмывают холодным изотоническим раствором хлорида натрия, а затем помещают в кислый ацетатный буфер, фракционируют и исследуют иммунохимически. В-лимфоциты-антителопродуценты синтезируют антитела и различаются по стадиям и направлению антигензависимой дифференцировки. Дифференцировка антителопродуцента В-лимфоцита связана со способностью к превращению в клон плазматических клеток, а также к смене класса синтезируемого антитела (с тем же идиотипом): синтез IgM сменяется синтезом IgG или IgА, а при локальном иммунном ответе синтез IgM сменяется синтезом IgE-антител. Не происходит смены классов Ig при переходе В-лимфоцита в плазматическую клетку. Плазматические клетки, синтезирующие IgM, – столь же зрелые по уровню дифференцировки, как и плазматические клетки, синтезирующие IgA или IgE. В-лимфоциты – антителопродуценты, синтезирующие IgM. Активный синтез IgM антител начинается уже в первые 2–3 дня после рождения под влиянием естественной антигенной стимуляции. К IgM-антителам принадлежат изогемагглютинины, холодовые агглютинины, ревматоидный фактор, высокоавидные бактерицидные антитела. IgM не проходит через плаценту, поэтому групповые и резус-изогемагглютинины не попадают от матери ребенку. В секретах IgM обнаруживается в небольшом количестве, он представлен главным образом в русле крови, где длительность его жизни составляет 5–9 дней. В-лимфоциты – антителопродуценты, синтезирующие IgG. В-лимфоциты играют основную роль во вторичном иммунном ответе. Это предшественники плазматических клеток, секретирующих IgG в сыворотку. Взаимосвязь синтеза IgG и IgM не только прямая, но и обратная – увеличение синтеза IgG, как правило, угнетает синтез IgM. Синтез IgG антител начинается на 1–4-м месяце после рождения и к 3-летнему возрасту достигает уровня синтеза взрослого. Выделяют 4 субкласса IgG. К цитофильным антителам относятся IgG1 и IgG3, которые в большом количестве прикрепляются к моноцитам. IgG способны связывать комплемент, а комплексы IgG-антиген реагируют с тромбоцитами, вызывая секрецию вазоактивных аминов. IgG-антитела в большом количестве находятся в сыворотке, легких, желудочно-кишечном тракте, печени. Молекулы IgG легко проходят через плаценту, создавая иммунитет у плода. В-лимфоциты и плазмоциты, синтезирующие IgG, находятся в селезенке и лимфатических узлах. Дефицит IgG обычно сочетается с дефицитом IgM. Такое сочетание приводит к атрофии лимфатических узлов, отсутствию в них плазмоцитов, а клинически напоминает агаммаглобулинемию. В-лимфоциты – антителопродуценты, синтезирующие IgА. В-лимфоциты, несущие IgA на мембране, служат предшественниками плазматических клеток, синтезирующих IgА. IgA обладает выраженной активностью против вирусных, бактериальных, паразитарных и алиментарных антигенов, выполняет функцию местной защиты всех слизистых оболочек. Если в сыворотке крови IgA в норме значительно меньше, чем IgG, то в секретах IgA превышает уровень IgG в 100–1000 раз. IgA оказывает прямое бактерицидное действие без участия комплемента. Дефицит IgA вплоть до полного его отсутствия встречается нередко (1 : 700) и представляет собой наиболее частую форму иммунодефицитности. Дефицит IgA приводит к злокачественному течению полиомиелита, эпидемического паротита. При дефиците IgA наблюдаются рецидивирующие инфекции. В-лимфоциты и плазматические клетки, синтезирующие IgА, расположены в лимфатической ткани под слизистыми оболочками. В тканях их больше, чем в периферической крови, более чем в 6 раз. Дифференцировка и созревание В-клеток особенно чувствительны к регулирующему влиянию Т-лимфоцитов. При удалении вилочковой железы даже у взрослого уровень IgA довольно быстро снижается, в том числе уменьшается выделение секреторного IgA. Уровень IgM при этом практически не меняется. При возрастной инволюции вилочковой железы также уменьшается содержание IgA. В тканях кишечника при дефиците В-лимфоцитов может происходить накопление IgM В-лимфоцитов. Дефицит IgA может проявляться в различных формах в зависимости от уровня поражения. При нарушении синтеза мономера наблюдается дефицит сывороточного и секреторного IgA.. Клинически при этом определяется компенсаторное увеличение вилочковой железы и периферических лимфатических органов, особенно лимфатических узлов пищеварительного тракта. При отсутствии секреторного IgA наблюдается предрасположение к различным формам локальных поражений желудочно-кишечного тракта (таким как муковисцидоз, язвенный колит, терминальное воспаление подвздошной кишки, стоматиты), а также ревматоидному артриту и другим заболеваниям соединительной ткани. Большое значение имеет дефицит IgA в механизме развития аутоиммунных заболеваний, болезнях иммунных комплексов, болезни тяжелых цепей. В-лимфоциты, синтезирующие IgE. IgE В-лимфоциты при обычных исследованиях не выделяются ввиду их малого количества. Количество В-лимфоцитов, синтезирующих IgE, увеличивается при аллергических состояниях и паразитозах, что было выявлено еще в 1983 г. Бережной Н. М. и Ялкут С. И. Эти антитела способны к фиксации на поверхности тучных клеток и базофилов, имеющих специальный рецептор. Фиксированные на поверхности этих клеток IgE антитела взаимодействуют с антигеном, вызывая при этом дегрануляцию тучных клеток и базофилов и выход из них субстанций анафилаксии. Способность IgE после взаимодействия с антигеном «запускать» реакцию гиперчувствительности немедленного типа определила специальное название антител класса IgE – «реагины». IgE В-лимфоциты расположены главным образом в подслизистых слоях дыхательного и пищеварительного трактов, а также в коже и ближайших к покровным тканям лимфатических узлах. Основная функция IgE В-лимфоцитов заключается в синтезе антител местной защиты в ответ на проникновение малого количества антигена, поэтому синтез IgE имеет автономную регуляцию, которую осуществляют специальные Т-хелперы и Т-супрессоры, способствуя переключению синтеза IgM на IgE и соответствующей этому дифференцировке В-лимфоцитов. Ответ на антиген с помощью IgE антител происходит без участия лимфоцитов ближайших и отдаленных лимфатических узлов. Запуск местной воспалительной реакции завершается клетками местной защиты – базофилами и эозинофилами. Таким образом, в осуществлении гомеостаза IgE выполняет барьерную функцию. Содержание IgE в циркуляции увеличивается при патологических процессах с проявлениями гиперчувствительности немедленного типа. Кроме того, содержание IgE увеличивается при ряде паразитозов и вирусных инфекций. Дефицит IgE отмечен при синдроме Луи – Барр (нарушение координации движений – расширение сосудов кожи и слизистых оболочек); при этом заболевании снижено количество IgE и IgA, в связи с чем угнетены аллергические реакции и функция местной защиты. Противоположная картина наблюдается при синдроме Вискотта – Олдрича: увеличение IgA и IgE в сыворотке и тканевых жидкостях на фоне снижения IgM и клеток иммунологической памяти. Резкое увеличение содержания IgE отмечено при изолированном дефиците IgА. Гуморальные медиаторы В-лимфоцитов Антитела, синтезируемые антителопродуцирующими В-лимфоцитами и плазматическими клетками, составляют первую гуморальную систему иммунной защиты организма. Следует отметить, что, кроме важной роли в осуществлении специфической гуморальной защиты, иммунологлобулины участвуют в клеточных реакциях, прикрепляясь к рецепторам лимфоцитов, макрофагов, тучных клеток, базофилов. Участие В-лимфоцитов в выработке гуморальных медиаторов (вторая гуморальная система иммунной защиты) связано со способностью к секреции ряда лимфокинов. К ним относятся стимулятор В-клеток, митогенный фактор В-клеток, хелперный фактор, выделяемый В-лимфоцитами костного мозга, супрессорный фактор В-лимфоцитов костного мозга, супрессорный фактор, выделяемый более зрелыми В-лимфоцитами. Фактор торможения миграции макрофагов секретируется В-лимфоцитами даже в большем количестве, чем Т-лимфоцитами. Плазматические клетки синтезируют гуморальный фактор, мобилизирующий ионы Са++. В-лимфоциты-супрессоры – строго специфичные по отношению к антигену клетки, на поверхности которых определяются молекулы IgG. Эффект супрессии проявляется только к однородным по тканевой совместимости клеткам и направлен против хелперов, киллеров и активированных макрофагов. В-супрессоры расположены главным образом в костном мозге и селезенке, при активации они пролиферируют и продуцируют антитела. Определенная роль принадлежит В-супрессорам в создании устойчивости к трансплантатам у новорожденных. Антигеннеспецифическая супрессия В-лимфоцитов костного мозга, напротив, не ограничена барьером гистосовместимости и, вероятно, связана с синтезом и секрецией неспецифического супрессорного фактора. В-лимфоциты иммунологической памяти имеют на мембране комплекс антиген – антитело, прикрепляющийся к Fc-рецептору клетки. Они активируются при вторичном иммунном ответе и пролиферируют с образованием клона плазматических клеток, синтезирующих Ig того же класса, что и клетка иммунологической памяти. Цитотоксические В-лимфоциты (киллеры, так называемые К-клетки) отличаются от прочих В-лимфоцитов отсутствием поверхностных Ig. Цитотоксическая функция К-клеток – антителозависимая и связана с прикреплением к Fc-рецептору В-лимфоцитов цитотоксических антител. К-клетки находятся в конкурентных отношениях с блокирующими антителами, а именно не дающими достаточного цитотоксического эффекта. Соединяясь с антигенами, клетки-мишени, блокирующие антитела, делают ее недоступной для действия киллеров всех видов. К-клетка, присоединяя к своей поверхности большое количество цитотоксических антител, способна повреждать клетку-мишень. Направленность специфического иммунитета в каждом конкретном случае во многом зависит от соотношения между содержанием К-клеток и блокирующих антител. Дифференцировка В-лимфоцитов Неоднородность В-лимфоцитов и их субпопуляций, а также выделение в последние годы еще недостаточно изученных субпопуляций (В-супрессоров, В-клетки-киллеров) не позволяют представить достаточно достоверную последовательность дифференцировки В-клеток. Ни Т– ни В-лимфоидные клетки Лимфоидные клетки, не имеющие Т– и В-маркеров, представляют собой оставшуюся после выделения Т– и В-клеток субпопуляцию. В ее состав входят стволовые клетки костного мозга, являющиеся предшественниками В-, Т– или обеих субпопуляций лимфоцитов, естественные киллеры. Нередко к этой субпопуляции относят В-клетки-киллеры. Несмотря на немногочисленное представительство этой субпопуляции в периферической крови (не более 5–10% среди лимфоцитов), все входящие в нее группы клеток имеют большое значение для кроветворения и иммунного ответа. Глава 6. Эритроциты Кроветворение плода происходит сперва в желточном мешке, стебле хориона, затем главным образом в печени и наконец по большей части в костном мозге. Впервые эритроциты обнаруживаются у 19-дневного человеческого эмбриона в кровяных островках желточного мешка. Поскольку периферические клетки желточного мешка образуют сосуды, а центральные – примитивные клетки крови, создается впечатление, что последние возникают внутрисосудисто, и о первом этапе кровообразования говорят как о внутрисосудистом. К 22-му дню эмбрионального периода первые кровяные клетки проникают в мезодермальную ткань эмбриона, в сердце, аорту, артерии, но большая их часть находится в желточном мешке. На 6-й неделе снижается активность кроветворения в желточном мешке. Первый период кроветворения (преимущественно образования эритроцитов) полностью заканчивается к началу 4-го месяца внутриутробного периода. Примитивные кроветворные клетки желточного мешка рано подвергаются дифференцировке, накапливая гемоглобин и превращаясь в примитивные эритробласты. П. Эрлих назвал примитивные эритробласты мегалобластами, которые представляют собой большие по диаметру клетки (от 9 до 30 мкм), овальной формы с ядром и цитоплазмой разной степени зрелости. Внешне мегалобласты эмбриона напоминают мегалобласты при пернициозной анемии, хотя дефицита витамина В12 у плода не отмечается. В тот момент, когда эмбрион достигает в длину 7,5–8 мм, до 15% его мегалобластов теряют ядро и вымываются в кровь в виде мегалоцитов, также в циркуляции встречаются ретикулоциты. К 3-му месяцу внутриутробной жизни число ретикулоцитов в кровотоке уменьшается, преобладают зрелые эритроциты первой генерации. Считается, что эритроциты первой генерации к началу месяца полностью исчезают из кровотока, не переходя в следующую клеточную генерацию. Число ретикулоцитов вновь возрастает к этому времени. После 5 недель внутриутробного развития начинается второй период кроветворения, который носит название печеночного. Кроветворение в печени достигает максимума к 5-му месяцу. Кроветворение второго периода преимущественно эритроидное, хотя с 12-й недели в печени циркулирует много клеток – предшественниц не только красного ряда, но и гранулоцитарно-макрофагальных. In vitro (в пробирке) они образуют макрофагальные и нейтрофильные колонии. После 20 недель интенсивность эритропоэза в печени снижается. Эритропоэз в печени также происходит внесосудисто. Кроветворные клетки располагаются между клетками печеночной паренхимы, а также в околосинусных пространствах эмбриональной печени. На 3-м месяце эмбрионального периода в эритропоэз включается селезенка, но у человека ее роль в пренатальном кроветворении весьма ограничена. Печеночный период эритропозэа определяется как нормобластический. Эритроциты этой генерации большей частью круглые. На 4–5-м месяце внутриутробной жизни начинается миелоидный период кроветворения, который постепенно вытесняет гепатолиенальный. К моменту рождения у доношенного плода экстрамедуллярные очаги эритропоэза почти полностью ликвидируются (единичные остаются в печени), а костный мозг развит полностью. Медуллярный эритропоэз также совершается экстраваскулярно (вне кровеносных сосудов), в строме костного мозга. Миелоидный эритропоэз плода мало отличается от эритропоэза взрослого. Общими закономерностями эмбрионального эритропоэза являются постепенное уменьшение диаметра и объема эритроцитов и увеличение их числа. Однако даже эритроциты и ретикулоциты новорожденного больше по размерам, чем у взрослого. Эритроциты плода живут меньше, чем эритроциты взрослого человека, – 45–70 дней (в среднем 20–30 дней) вместо 120. Кроветворение плода имеет разную органную локализацию: кроветворение в желточном мешке сменяется печеночным, затем кроветворением в селезенке (параллельно печеночному) и наконец костномозговым. В какой-то момент кроветворение происходит в нескольких органах одновременно, и в циркуляции в одно и то же время может оказаться зрелое потомство из разных кроветворных органов. Смене кроветворных органов у плода отвечает смена типов продуцируемого красными клетками гемоглобина. Примитивные эритрокариоциты, поступающие из желточного мешка, продуцируют преимущественно эмбриональные гемоглобины. До 37-го дня преобладает продукция гемоглобинов данного типа. В печени потомство эритроидных клеток-предшественниц содержит в основном фетальный гемоглобин. До 30 недель внутриутробного периода у человеческого плода в циркуляции преобладает фетальный гемоглобин, хотя с 10 недель начинают появляться эритроциты с гемоглобином взрослого – HbА. HbA до 30 недель составляет лишь 10%. Остальное составляют преимущественно клетки с HbF; еще есть и следы эмбриональных гемоглобинов. После 30 недель содержание клеток с HbA начинает нарастать, что совпадает с развитием костномозгового кроветворения. К моменту рождения фетальный гемоглобин составляет только 49%, остальное приходится на HbА. Таким образом, красные клетки плода одновременно продуцируют несколько типов гемоглобина. Кроветворение новорожденных. В течение нескольких дней после рождения у новорожденного наблюдается увеличение количества эритроцитов: число эритроцитов от 4,5 ? 1012/л (в среднем к моменту рождения), через 12 ч после рождения повышается до 5,5 ? 1012/л, гемоглобин – от 156 г/л до 200 г/л. Спустя несколько дней у новорожденного снижается продукция эритропоэтина, что совпадает со снижением уровня эритроцитов и гемоглобина в крови. Такая картина наблюдается в течение первых l – 2 месяцев жизни. Между 60-м и 90-м днем жизни вновь повышается содержание эритропоэтина, нарастает ретикулоцитоз, нормализуется эритропоэз. В течение первого года жизни изменяется антигенная структура эритроцитов: антиген i, преобладающий на эритроцитах новорожденного, сменяется антигеном I. Смена антигенов эритроцита совпадает с исчезновением так называемых фетальных эритроцитов – клеток, содержащих фетальный гемоглобин. К первому году жизни фетальный гемоглобин уже не превышает 1%. Форма и размеры эритроцита. Эритроцит человека в норме имеет двояковогнутую, дискоидную форму. Считается, что плоский диск лучше всего адаптирован к транспорту веществ из клетки и внутрь ее и к диффузии газов к центру клетки. В настоящий момент доказано, что двояковогнутая форма обладает незначительными диффузионными преимуществами. Однако объем, соответствующий диску, имеет в 1,7 раза большую поверхность, чем такой же объем, соответствующий сфере, и может умеренно изменяться без растяжения мембраны клетки. Двояковогнутая форма эритроцита, эластичность, деформируемость и сохранение структуры клетки при удалении из нее гемоглобина, когда остается так называемая тень эритроцита, заметно зависят от особенностей его строения, прежде всего скелета клетки. Структура плазменной мембраны у эритроцита такая же, как у ядерных клеток, но его цитоскелет отличается от цитоскелета этих клеток. Этот цитоскелет называют еще скелетом мембраны, и по его расположению и по тому, что он придает прочность по большей части мембране, обеспечивается единство ее липидного слоя, в то же время придавая мембране внутреннюю подвижность и гибкость. С помощью электрофореза в мембране и цитоскелете эритроцита выделено 8 типов белка. Основным опорным стабилизирующим белком цитоскелета эритроцита является спектрин – специфический белок эритроцита, хотя одна из его важных функций по соединению с актином (водорастворимый глобулярный белок) и актина с мембраной клетки присуща и некоторым белкам других клеток. С изменениями цитоскелета эритроцита связаны некоторые формы гемолитических анемий – наследственные эллиптоцитоз, пиропойкилоцитоз, отдельные варианты наследственного микросфероцитоза. Цитоскелет эритроцита играет важную роль в его способности к деформации. Дисковидный эритроцит может легко пройти фильтр с отверстием 3 мкм. За 100–120 дней циркуляции в организме способность эритроцита к деформации снижается. С возрастом снижается стойкость эритроцитов к осмотическому разрушению эритроцитов, к саморазрушению, в меньшей степени – к механической травме. Стареющие сферические эритроциты, как и сфероциты при патологии, имеющие пониженную способность к деформации, не могут проходить через миллипоровые фильтры 3 мкм, задерживаются они и селезенкой. Возможно, снижение деформируемости с возрастом эритроцита и сферуляция клетки связаны с изменением цитоскелета. У старого эритроцита обнаруживается агрегация спектрина и гемоглобина. В деформируемости эритроцита играет роль не только цитоскелет, но и липиды мембраны, в частности соотношение фосфолипидов и холестерина в мембране, которое определяет текучесть мембраны у всех клеток вообще. Это свойство также может иметь отношение к стойкости мембраны эритроцита. Текучесть клеточных мембран меняется при их отмывании. Размеры нормального эритроцита человека изменчивы, но можно установить пределы средних колебаний. Их цифровое значение зависит от методов определения (например, измеряется ли диаметр эритроцита в мазке или в плазме). Существуют некоторые колебания, связанные с полом. Эритроциты у новорожденных больше по размеру и объему, чем у взрослых. Диаметр нормального эритроцита человека – 7,5–8,3 мкм. Он несколько уменьшается с возрастом клетки. Толщина эритроцита – 2,1 мкм, средний объем – 86,1 мкм3, а площадь поверхности – 145 мкм2. После потери нормоцитом ядра он превращается в ретикулоцит. Ретикулоцит пребывает в костном мозге 30–40 ч. Костномозговые ретикулоциты, каково бы ни было их абсолютное количество, образуют небольшой резерв красной крови. Постоянство процента ретикулоцитов крови в норме позволяет судить об интенсивности кроветворения. Выход эритроцитов из костного мозга регулируется гуморально, в частности селезенкой и эритропоэтином (гормон, стимулирующий образование эритроцитов в костном мозге). В норме время созревания ретикулоцита в периферической крови составляет 35–45 ч. В случае выхода в кровь резервных ретикулоцитов это время иногда удлиняется, так как поступают ретикулоциты первых стадий созревания. Содержание ретикулоцитов в крови в норме, по данным разных авторов, составляет от 0,8–1,3 до 0,2–2%. При исследовании методом электронной микроскопии ретикулоцит имеет вид клетки неправильной формы с остатками органелл. Маленькие митохондрии и центриоли сконцентрированы в той области клетки, где на стадии нормоцита отделялось ядро. Рибосомы рассеяны по цитоплазме. При созревании митохондрии уменьшаются в числе и размерах, полирибосомы превращаются в монорибосомы. По мере того как ретикулоцит самопереваривает органеллы, в нем появляются вакуоли. Обмен железа Железо является одним из основных по значению микроэлементов организма. Почти все железо входит в состав различных белков. Из них наиболее важен гемоглобин, функция которого – перенос кислорода от легких к тканям. Гемоглобин состоит из небелковой части – гема, и белковой части – глобина. В молекуле гема железо связано с протопорфирином. Гем не только входит в состав гемоглобина, он содержится в миоглобине, цитохромах, входит в состав каталазы, лактопероксидазы. Основной белок, содержащий железо и не имеющий гемовой группы, – ферритин. Он содержит железо запасов. Железо входит и в состав производного ферритина – гемосидерина. Не содержит группы гема белок трансферрин, переносящий железо. Железо в негемовой форме есть в ряде ферментов (аконитазе, ксантиноксидазе). Основное количество железа в организме (57,6%) входит в состав гемоглобина и содержится в эритроцитах. Значительное количество железа есть в мышцах (27,6%). Большая часть этого железа входит в состав ферритина (68,1% железа мышц), остальная часть включена в миоглобин (21,9%). В печени откладывается 7,8% железа организма. Железо печени в основном входит в состав ферритина и гемосидерина. Трансферрин – белок плазмы крови, относящийся к глобулинам. Он имеет 2 активных участка, каждый из которых может связать по одному атому железа в трехвалентной форме. Основной синтез трансферрина у людей происходит в печени. За сутки производится 12–24 мг трансферрина на 1 кг массы, т. е. 5–9% всего количества этого белка. Содержание железа в организме зависит в основном от его всасывания. Выделение железа из организма – процесс, недостаточно регулируемый. Существует сложный механизм, препятствующий всасыванию избыточного количества железа. Хотя теоретически весь кишечник, включая толстую кишку, способен всасывать железо, основное количество железа всасывается в двенадцатиперстной кишке, а также в начальной части тощей кишки. Чем больше дефицит железа, тем дальше в тощую кишку распространяется зона его всасывания. Процесс всасывания железа у человека включает в себя проникновение железа в слизистую оболочку из просвета кишки, проникновение железа из слизистой оболочки в плазму, заполнение запасов железа в слизистой оболочке и влияние этих запасов на всасывание. Железо проникает в слизистую оболочку из просвета кишки всегда быстрее, чем поступает из слизистой оболочки в плазму. Хотя обе величины зависят от потребностей организма в железе, проникновение железа в слизистую оболочку кишки меньше зависит от содержания железа в организме, чем проникновение железа из слизистой оболочки в плазму. При повышенной потребности организма в железе скорость его поступления в плазму из слизистой оболочки приближается к скорости проникновения в слизистую оболочку кишки. Железо при этом в кишке практически не откладывается. Прохождение железа через слизистую оболочку занимает несколько часов. В этот период кишка невосприимчива к дальнейшему всасыванию железа. Через некоторое время железо вновь всасывается с такой же интенсивностью. При уменьшении потребности организма в железе замедляется его проникновение в кишку, еще больше уменьшается поступление железа из слизистой оболочки в плазму. Большая часть железа, которое не всасывается, откладывается в кишке в виде ферритина. Всасывание железа, входящего в состав гема, происходит значительно более интенсивно, чем всасывание неорганического пищевого железа. В слизистой оболочке кишки имеется фермент гемоксигеназа, необходимый для распада молекулы гема на билирубин, окись углерода и ионизированное железо. При нормальном содержании железа в организме значительная его часть проходит через слизистую оболочку кишки в ток крови, а определенная часть задерживается в стенке кишки. При сидеропении в слизистой оболочке задерживается значительно меньшая часть, основная часть железа оказывается в плазме. При избытке железа в организме основная часть железа, проникшего в слизистую оболочку, в ней и задерживается. Впоследствии эпителиальная клетка, наполненная железом, движется от основания к концу ворсинки, затем слущивается и выводится с калом вместе с невсосавшимся железом. Этот физиологический механизм всасывания действует при обычных содержащихся в нормальной пище концентрациях железа. Если концентрация железа превышает в десятки и сотни раз физиологическую, то всасывание ионного двухвалентного железа во много раз возрастает. Это следует учитывать при лечении больных солями двухвалентного железа. Трехвалентное железо практически не всасывается ни в физиологических концентрациях, ни в избыточных. Всасывание пищевого железа строго лимитировано: за сутки всасывается не более 2–2,5 мг. Железо содержится во многих продуктах как растительного, так и животного происхождения. Высока концентрация железа в мясе, печени, почках, много железа содержат бобы сои, петрушка, горох, шпинат, сушеные абрикосы, чернослив, изюм. Значительное количество железа содержится в рисе, хлебе, яблоках. Однако имеет значение не количество железа в продукте, а его всасывание из данного продукта. Из продуктов растительного происхождения железо всасывается очень ограниченно, в значительно большей степени – из большинства животных продуктов. Железо, входящее в состав белков, содержащих гем, всасывается значительно лучше, чем из ферритина и гемосидерина, а железо из печени всасывается значительно меньше, чем из мяса. Поэтому хуже всасывается железо из рыбы, так как в ней железо присутствует в основном в виде гемосидерина и ферритина, а в телятине до 90% железа содержится в виде гема. На всасывание железа влияет ряд факторов. Частота сочетания железодефицитной анемии с ахилией (отсутствие соляной кислоты и фермента пепсина в желудочном соке) еще в XX в. дала основание предполагать, что железо всасывается лишь при нормальной желудочной секреции и ахилия является одним из основных факторов, приводящих к развитию железодефицитной анемии. Однако исследования показали, что нормальная желудочная секреция влияет на всасывание некоторых форм железа, однако это не главный фактор в регуляции его всасывания. Хлористоводородная кислота влияет лишь на всасывание трехвалентного железа. Желудочная секреция не влияет на всасывание железа, входящего в состав гема. В норме всасывание гемоглобинового железа у здоровых женщин в среднем составляет 16,9 ± 1,6%, у мужчин – 13,6 ± 1%. При железодефицитной анемии всасывание железа резко повышено и не различается у лиц с нормальной и пониженной секрецией. Нормальным оказалось всасывание железа у лиц, перенесших удаление части желудка. У лиц с атрофическим гастритом без анемии всасывание гемоглобинового железа не отличалось от всасывания железа у здоровых лиц. Доказано, что оксалаты, фитаты, фосфаты входят в комплекс с железом и снижают его всасывание, а ряд веществ усиливает всасывание железа. К ним относятся аскорбиновая, янтарная, пировиноградная кислоты, фруктоза, сорбит. Всасывание железа усиливается под влиянием алкоголя. Недостаток кислорода, снижение запасов железа в организме, активизация кроветворения усиливают всасывание железа. Влияют на всасывание железа насыщение трансферрина, концентрация железа плазмы, скорость оборота железа, уровень эритропоэтина. После всасывания железо связывается с трансферрином, который переносит железо к эритрокариоцитам костного мозга. Кроме того, трансферрин переносит железо от клеток, где хранятся его запасы, к эритрокариоцитам, а также от фагоцитирующих макрофагов, где железо распадается, к клеткам костного мозга и к местам, где сохраняются запасы железа. Одна молекула трансферрина присоединяет 2 атома железа. На мембране эритрокариоцита и мембране ретикулоцитов наблюдаются специфические участки для обратимого присоединения трансферрина. Связывание железа с трансферрином и его освобождение – это активные процессы, которые подавляются ингибиторами ферментов. К поверхности ретикулоцита могут присоединяться 25 000–50 000 молекул трансферрина, нагруженных железом. Меченый по железу трансферрин легко присоединяется к ретикулоцитам, но не присоединяется к лейкоцитам, тромбоцитам и зрелым эритроцитам. После того как трансферрин «разгружает» железо на поверхности эритрокариоцитов, оно проникает внутрь клетки. Трансферрин в большинстве случаев способен возвращаться в плазму, но некоторые его молекулы проникают внутрь эритрокариоцита и связываются с молекулой носителя. Железо проникает в митохондрии, где происходит синтез гема из протопорфирина и железа. Образование ферритина происходит в эритрокариоците из белка апоферритина, синтезируемого в клетке, и железа, проникшего в клетку. Наиболее вероятно, что синтез ферритина в эритрокариоците нужен для удаления из клетки избыточного железа, не вошедшего в гемоглобин. Этот ферритин собирается в лизосомах, а затем удаляется из клетки как в костном мозге, так и в циркуляции после удаления из клетки ядра. В удалении гранул железа из циркулирующей клетки участвует, по-видимому, селезенка, так как в эритроцитах людей после удаления селезенки обнаруживаются гранулы железа, а в норме выявить их в зрелых эритроцитах не удается. Основным белком, используемым для сохранения избытка железа в организме, является ферритин. Ферритин – это водорастворимый комплекс гидроокиси трехвалентного железа и белка – апоферритина. Гидроокись железа соединена с остатком фосфорной кислоты. Молекула ферритина напоминает по форме грецкий орех: скорлупа ореха – это белок апоферритин, а внутри находятся в различном количестве атомы железа, почти вплотную прилегающие один к другому. Ферритин может вместить до 4500 атомов железа, практически 1 молекула содержит около 3000 атомов. Молекулярная масса ферритина зависит от числа атомов железа, а этот показатель может колебаться. В среднем молекулярная масса ферритина близка к 460 000. Ферритин в норме имеется в плазме и практически почти во всех клетках организма, но больше всего – в печени и мышцах. Гемосидерин – белок, содержащий железо, обнаруживаемый в фагоцитирующих макрофагах и их производных, в макрофагах костного мозга и селезенки, в купферовских клетках печени. Гемосидерин – это частично денатурированный и депротеинизированный ферритин. Иммунологически гемосидерин полностью идентичен ферритину. Молекула ферритина содержит 20% железа, а в гемосидерине железа больше – 25–30%. В отличие от ферритина гемосидерин нерастворим в воде. Как гемосидерин, так и ферритин используются в качестве белков запаса, однако скорость мобилизации гемосидерина значительно более медленная, чем ферритина. Железо запасов может быть как в паренхиматозных клетках, так и в фагоцитирующих макрофагах. В норме основную часть железа, связанного с трансферрином, организм использует для кроветворения. Фагоцитирующие макрофаги, получившие железо при разрушении в них эритроцитов, в основном передают это железо трансферрину, который вновь использует его для кроветворения. Паренхиматозные клетки тоже содержат железо, но в основном в запасах, и лишь малая часть его передается трансферрину и используется для эритропоэза. Паренхиматозные клетки в свою очередь получают железо из трансферрина. В отличие от железа макрофагов железо, находящееся в паренхиматозных клетках, расходуется медленно. Аскорбиновая кислота увеличивает освобождение железа из макрофагов, но не влияет на его освобождение из паренхиматозных клеток. Освобождение железа из паренхиматозных клеток увеличивается при кровотечениях и уменьшается при массивных гемотрансфузиях. При кровотечениях уменьшается захват эритроцитов макрофагами, следовательно, освобождение железа макрофагами в такой ситуации имеет меньшее значение. Понятие «лабильный пул железа» появилось при изучении кинетики железа. Оно покидает плазму и входит в интерстициальное пространство тканей. Там железо может связываться с клеточными мембранами. Его часть возвращается в плазму, и этот процесс приводит к отклонению линии клиренса железа, что выявляется в 1-й или во 2-й день после введения радиоактивного железа. Изменение в наклоне линии зависит от количества так называемого лабильного пула. Рассчитано, что в норме лабильный пул содержит 80–90 мг железа. Тканевое железо – это 6–8 мг железа, входящего в состав цитохромов и других ферментов всех тканей организма. Мужчины за сутки теряют около 1 мг железа. Потери железа у неменструирующих женщин соответствуют этим цифрам. Потери железа у менструирующих женщин намного превышают потери железа у мужчин. Они слагаются из потерь, свойственных мужчинам, и потерь, свойственных только женщинам: потери железа во время менструальных кровотечений, во время беременности, родов и лактации. По данным различных исследований, потери железа у здоровых женщин колеблются от 2 до 79 мг за одну менструацию. В среднем они теряют за время менструации 30 мл крови, что соответствует 15 мг железа, однако у 11% здоровых женщин количество теряемой крови превышает 80 мл (40 мг железа). Такую кровопотерю гинекологи считают нормальной. У рожавших женщин кровопотеря несколько больше, чем у нерожавших. Таким образом, при расчете потери железа на 1 день месяца следует учитывать, что при нормальных менструациях женщины теряют в день от 0,5 до 1,2 мг железа. Во время беременности потеря железа составляет не менее 700–800 мг, а потребности в железе во время беременности большие, они составляют 800–1200 мг. Глава 7. Система гемостаза Система гемостаза – биологическая система, благодаря которой обеспечивается, с одной стороны, сохранение жидкого состояния крови, а с другой – предупреждение и остановка кровотечений путем поддержания структурной целостности стенок кровеносных сосудов и достаточно быстрого тромбирования последних при повреждениях. Важность данной системы для сохранения жизнеспособности организма определяется тем, что она препятствует убыли крови из циркуляторного русла и тем самым способствует обеспечению нормального кровоснабжения органов, сохранению необходимого объема циркулирующей крови. Гемостаз реализуется в основном тремя взаимодействующими между собой функционально-структурными компонентами – стенками кровеносных сосудов (в первую очередь их интимой), клетками крови и плазменными ферментными системами – свертывающей, фибринолитической (плазминовой), калликреин-кининовой. Система подчинена сложной нейрогуморальной регуляции, в ней четко функционируют механизмы положительной и отрицательной обратной связи, вследствие чего клеточный гемостаз и свертывание крови вначале подвергаются самоактивации, дальнейшая регуляция связана с нарастанием антитромботического потенциала крови. Вышеперечисленные механизмы создают необходимые условия для самоограничения процесса свертывания, в связи с чем локальная активация системы в местах тромбообразования не трансформируется при правильном функционировании указанных механизмов во всеобщее свертывание крови систему гемостаза. В осуществлении гемостаза немаловажное значение имеет не только собственно свертывающая система крови. Наряду с ней также реагируют и сами сосуды (спазм, открытие шунтов выше места повреждения) и клетки крови – тромбоциты и отчасти эритроциты. Также следует выделить, что тромбоцитам, а не именно свертыванию крови принадлежит ведущая роль в первичной остановке кровотечений из микрососудов (диаметром до 100 мкм), наиболее ранимых и чаще всего являющихся источником геморрагии. Время кровотечения из мельчайших сосудов кожи, определяемое по Дьюку или другими способами, всегда удлинено при тромбоцитопениях и тяжелых дисфункциях кровяных пластинок и остается нормальным при гемофилиях и многих других нарушениях свертываемости крови. Вследствие этих причин сосудисто-тромбоцитарная реакция на потерю крови часто обозначается как начальный, или первичный, гемостаз, а свертывание крови – как вторичная гемостатическая реакция, хотя оба эти механизма включаются не строго последовательно друг за другом, а на значительном отрезке времени функционируют одновременно и сопряженно. Клеточный гемостаз в эволюционном отношении является более ранним и в определенной степени родоначальным механизмом. Так, у реликтовых низших беспозвоночных остановка кровотечений обеспечивается только клетками гемолимфы, и в плазме этих животных еще нет факторов свертывания. У более высокоорганизованных животных (омаров) в плазме уже появляется аналог фибриногена, но еще нет тромбина, и примитивное свертывание при удалении клеток крови идет под влиянием трансглутаминазы. И лишь у позвоночных свертывающая система плазмы получает высокое развитие и значительную автономию, хотя и у них выход из клеток активаторов свертывания играет важную роль в осуществлении гемостаза. Сосудисто-тромбоцитарный гемостаз Стенки кровеносных сосудов играют чрезвычайно важную роль не только в обеспечении гемостаза, но и в поддержании жидкого состояния крови. Интима сосудов, эндотелий обладают очень высокой тромбоустойчивостью, в силу чего сохранность этой внутренней выстилки – важнейшее условие сохранения жидкого состояния крови. В основе этой тромбоустойчивости лежат сложные и пока далеко не полностью расшифрованные механизмы (от отрицательного заряда цитоплазматической мембраны эндотелиальных клеток до их способности вырабатывать и секретировать вещества, препятствующие агрегации тромбоцитов, свертыванию крови, а также активаторы фибринолиза). Среди этих механизмов достаточно хорошо изучены следующие:
Вместе с тем в эндотелии синтезируются и факторы, необходимые для реализации гемостатических реакций. Так, например, маркером эндотелиальных клеток является фактор Виллебранда (антиген фактора VIII), необходимый для нормального прилипания тромбоцитов к коллагену и формирования тромбоцитарной пробки. В субэндотелиальном слое преобладают стимуляторы гемостаза, среди которых наиболее мощным агентом является коллаген, стимулирующий как прилипание тромбоцитов, так и внутренний механизм свертывания крови (активацию фактора XII). В субэндотелии содержатся и антитромботические активности. Для примера: гладкомышечные клетки, как и эндотелиальные, способны образовывать простациклин. Кроме того, они вырабатывают протеогликаны и среди них мощные ингибиторы свертывания крови и адгезии тромбоцитов. Коллаген реализует запуск не только свертывания крови, но и фибринолитической системы. Участие тромбоцитов в гемостазе определяется в основном следующими функциями этих клеток:
Ангиотрофическая функция. Тромбоцитам принадлежит важная роль в поддержании нормальной резистентности и функции микрососудов. С помощью электронной микроскопии и микроавторадиографии установлено, что тромбоциты периодически смыкаются с эндотелиальными клетками и «изливают» в них свое содержимое. Этот процесс поглощения кровяных пластинок эндотелиальными клетками идет особенно интенсивно после глубокой тромбоцитопении. В подобных условиях уже через 30 мин после переливания меченых тромбоцитов около 80% их массы оказываются в эндотелии. Из этого сделан вывод, что тромбоциты являются физиологическими «кормильцами» эндотелия и он не в состоянии извлекать ряд необходимых веществ прямо из плазмы. В нормальных условиях (без тромбоцитопении) эндотелий поглощает в среднем 35 000 кровяных пластинок из каждого микролитра крови за сутки. Следовательно, на ангиотрофическую функцию расходуется ежедневно около 15% всех циркулирующих в крови тромбоцитов. Если эндотелиальные клетки лишаются тромбоцитарной «подкормки», то они быстро подвергаются дистрофии и начинают пропускать через свою цитоплазму эритроциты. Проникновение эритроцитов происходит очень быстро – в течение нескольких минут и с большей энергией, о чем можно судить хотя бы по тому, что эритроцит, встретивший на своем пути ядро эндотелиальной клетки, либо отжимает его в сторону, либо ломает надвое. Вышедшие из капилляров эритроциты образуют мелкие кровоизлияния. Часть из них попадает в лимфу и через грудной лимфатический проток возвращается в систему кровообращения. При всех тромбоцитопениях содержание эритроцитов в лимфе грудного протока повышено, причем тем больше, чем значительнее дефицит кровяных пластинок. Таким образом, кровоточивость при тромбоцитопениях связана как с повышенной ломкостью микрососудов, так и с их повышенной проницаемостью для эритроцитов и других компонентов крови. Адгезивно-агрегационная функция. Способность тромбоцитов приклеиваться к поврежденным участкам сосудистой стенки и быстро образовывать в таких местах тромбоцитарную пробку, останавливающую кровотечение, была выявлена еще в конце XX в. Формирование тромбоцитарной пробки начинается с прилипанием тромбоцитов к субэндотелиальным структурам сосудистой стенки (к базальной мембране). Коллаген – главный стимулятор этого процесса, хотя прилипания тромбоцитов могут вызывать и другие компоненты соединительной ткани. Еще до взаимодействия с оголенной базальной мембраной тромбоциты подвергаются сложной внутренней перестройке – меняют свою форму (плоскую дискоидную на сферическую), выбрасывают длинные нитчатые отростки-псевдоподии, приобретая способность прикрепляться как к соединительной ткани, так и друг к другу. Известно, что в кровотоке указанная перестройка тромбоцитов происходит до того, как они достигнут поврежденного участка сосуда, вследствие чего к сосуду они уже доставляются, будучи подготовлены к прилипанию и агрегации. Одновременно с этим в кровотоке интенсивно идет и другой процесс – склеивание тромбоцитов друг с другом, в результате чего образуются конгломераты, состоящие из 3–15–20 клеток, которые приклеиваются к первично адгезировавшим тромбоцитам. В результате гемостатическая пробка быстро увеличивается в объеме и через 1–3 мин полностью заполняет просвет кровоточащего сосуда. Прилипание и агрегация тромбоцитов – сложная биологическая реакция, требующая участия ряда внешних и внутренних, исходящих из самих тромбоцитов, стимуляторов, энергетических затрат, глубокой перестройки свойств кровяных пластинок. Важнейшим плазменным кофактором адгезии тромбоцитов к коллагену является синтезируемый в эндотелии и циркулирующий в крови гликопротеин – фактор Виллебранда. Тромбоциты способны накапливать этот фактор в своих гранулах и выделять его в окружающую среду при активации (дегрануляция, «реакция освобождения»). Агрегация тромбоцитов реализуется рядом включающихся сопряженно и последовательно стимуляторов (агонистов): коллаген, АДФ, арахидоновая кислота и ее производные, адреналин, тромбин. В первичном запуске агрегации ведущая роль принадлежит АДФ. Его первые небольшие количества поступают из поврежденной сосудистой стенки и эритроцитов, мацерирующихся в зоне гемостаза. Затем АДФ выделяют в окружающую среду сами первично адгезировавшие и активированные тромбоциты в процессе присущей этим клеткам «реакции освобождения». В результате вышеописанных процессов концентрация АДФ в зоне гемостаза быстро нарастает. И спустя уже 20 с после перерезки артериолы около 50% всего имеющегося в тромбоцитах АТФ превращается в АДФ. Сопряженно с АДФ из тромбоцитов выделяются содержащиеся в тех же гранулах другие стимуляторы агрегации – адреналин, серотонин. Однако особое значение имеет то, что в лабилизированных тромбоцитах активируются мембранные фосфолипазы, циклооксигеназа и тромбоксан-синтетеза, в результате чего образуются мощные стимуляторы агрегации – арахидоновая кислота и ее производные, в том числе наиболее активный агрегант этой группы – тромбоксан А2. Аналогичным образом в эндотелии и гладкомышечных клетках стенок кровеносных сосудов активируется образование эндоперекисей простагландинов, но на последнем этапе под влиянием фермента простациклин-синтетазы в них образуется и выделяется в кровь мощный ингибитор агрегации тромбоцитов и вазодилататор – простациклин. Таким образом, система простагландинов – один из важных регуляторов агрегационной функции тромбоцитов и их взаимодействия с сосудистой стенкой. Для клиницистов знакомство с этими механизмами имеет существенное значение, поскольку с нарушением образования аденилатциклазы или с ее блокадой связан ряд наследственных («аспириноподобный синдром») и приобретенных, в том числе лекарственных, тромбоцитопатий. Препараты, ингибирующие эту систему, используются в антитромботической терапии, хотя целесообразность применения некоторых из них весьма проблематична, поскольку они в равной степени подавляют как агрегацию тромбоцитов, так и образование в эндотелии антитромботического агента – простациклина. Тромбин – чрезвычайно сильный агрегирующий агент, завершающий «реакцию освобождения» внутрипластиночных факторов, укрепление фибрином тромбоцитарной пробки. Важно, что агрегацию он вызывает в дозах, значительно меньше тех, какие необходимы для свертывания крови. Формирование тромбоцитарной пробки опережает свертывание, хотя отдельные волокна фибрина все же обнаруживаются в ней и на ранних этапах агрегации. Взаимодействуя с мембранным гликопротеином V, тромбин формирует на тромбоцитах рецепторы к активированным плазменным факторам свертывания X и V. Закрепляясь на тромбоците, фактор Ха получает защиту от антикоагулянтного действия антитромбина III и гепарина, что играет важную роль в реализации локального свертывания крови в зоне тромбирования сосудов. В механизме тромбоцитарного гемостаза важным и вместе с тем очень уязвимым звеном является «реакция освобождения» гранул и содержащихся в них агентов, необходимых как для осуществления гемостаза, так и для репарации поврежденной сосудистой стенки. Без «реакции освобождения» процесс агрегации обрывается на начальном этапе и не завершается формированием полноценной тромбоцитарной пробки. Это нарушение часто наблюдается как при наследственных, так и при вторичных (симптоматических) тромбоцитопатиях. Схема 1 Агрегация тромбоцитов 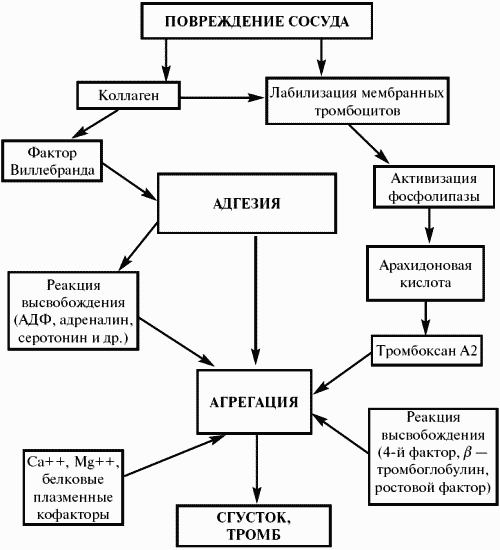 «Реакция освобождения» реализуется в 3 этапа:
В тромбоцитах различают 4 типа секретируемых гранул высокой электронно-оптической плотности. I тип – гранулы, с которыми секретируются важные для гемостаза небелковые компоненты: АТФ, АДФ, серотонин, пирофосфат, адреналин, кальций. II тип – гранулы, содержащие низкомолекулярные белки, фактор Виллебранда и фибриноген; наиболее важны в этих гранулах 2 разновидности пластиночного антигепаринового фактора (фактор 4 тромбоцитов, ПФ-4), ?-тромбоглобулин, ростовой (митогенный) фактор, стимулирующий синтез ДНК и деление клеток, фактор Виллебранда. III и IV тип гранул содержат ферменты, в основном кислые гидролазы; они секретируются позже и только под влиянием тромбина и коллагена, но не АДФ и адреналина. Агрегация тромбоцитов, особенно под влиянием АДФ, адреналина и тромбоксана, нуждается в ряде небелковых (ионы кальция и магния, фосфолипидный фактор) и белковых плазменных кофакторов. К последним относятся альбумин, термостабильный и термолабильный белковые кофакторы, фибриноген, некоторые компоненты глобулиновой фракции плазмы. Фибриноген необходим для агрегации в очень небольших количествах (немногим более 0,02 г/л), в связи с чем нарушения агрегации, связанные собственно с гипофибриногенемией, встречаются крайне редко. Все эти белки образуют вокруг тромбоцитов «плазматическую атмосферу», необходимую для полноценного функционирования этих клеток. Вместе с тем продукты ферментного расщепления белков (в частности, обусловленного плазмином расщепления фибриногена и фибрина) резко ингибируют агрегацию тромбоцитов. Таким же свойством обладают некоторые парапротеины и криоглобулины. Взаимодействие стимуляторов агрегации и ряда их плазменных кофакторов с кровяными пластинками происходит на мембране этих клеток – на предсуществующих или «открывающихся» в процессе активации рецепторах, в большинстве принадлежащих гликопротеинам (ГП). Многие виды патологии тромбоцитов, в том числе такие важные их формы, как тромбастения Гланцмана, макроцитарная тромбоцитодистрофия Бернара – Сулье, синдром Мей – Хегглина, обусловлены отсутствием или аномалией мембранных гликопротеиновых рецепторов. Гликопротеин I состоит из двух дисульфидносвязанных субъединиц – 1а или гликокалицина (молекулярная масса 130 000–160 000) и b (молекулярная масса 22 000). Первая является рецептором фактора Виллебранда; она необходима для прилипания тромбоцитов к субэндотелию (коллагену) и отчасти – для тромбинагрегации. Ее содержание в мембране тромбоцитов резко снижено при аномалии Бернара – Сулье. Гликопротеин II состоит из субъединиц IIа (молекулярная масса 110 000–130 000) и lib (молекулярная масса 23 000), необходим для всех видов агрегации тромбоцитов. Содержание резко снижено при тромбастении Гланцмана. Гликопротеин III, возможно, является вариантной формой гликопротеина II (молекулярная масса 114 000). Содержание в мембране снижено при тромбастении Гланцмана. Гликопротеин IV (молекулярная масса 85 000–100 000) отличается от других гликопротеинов резистентностью к трипсину и химотрипсину. Функция нуждается в уточнении. Гликопротеин V (молекулярная масса 68 000–89 000) является субстратом тромбина, которым селективно гидролизуется. Важен для реализации тромбин-агрегации. Тромбоцитарный гемостаз сам по себе вполне достаточен для полной остановки кровотечения в зоне микроциркуляции. Однако в более крупных сосудах с высоким кровяным давлением тромбоцитарная пробка, не укрепленная фибрином (без последующего свертывания крови), в лучшем случае лишь временно останавливает кровотечение, а затем часто не удерживается на месте, что ведет к его возобновлению. Влияние тромбоцитов на свертывание крови и фибринолиз. В тромбоцитах найдено много агентов, участвующих в свертывании крови. Однако многие из этих веществ являются не собственно тромбоцитарными факторами, а лишь адсорбированными тромбоцитами плазменными факторами свертывания. Плазма не только окружает тромбоциты снаружи, но омывает их изнутри, проникая вглубь клеток через ветвящиеся каналы. Они расширяются при активации кровяных пластинок. Многие факторы, участвующие в гемостазе и свертывании крови, сорбируются и концентрируются на поверхности тромбоцитов, другие накапливаются внутри клеток (в гранулах) и выделяются в процессе «реакции освобождения». Наконец, есть и такие компоненты, которые определяются в тромбоцитах в виде двух пулов – наружного (на мембране клетки) и внутреннего (чаще всего в гранулах). Следует отметить, что фибриноген, на долю которого приходится 3–4% всего белка плазмы, в тромбоцитах составляет 10–12% белка, причем 1/4 этого количества содержится в плотных гранулах II типа и секретируется при «реакции освобождения», а 3/4 – на оболочках кровяных пластинок. Точно так же фибринстабилизирующий фактор (фактор XIII) и фактор Виллебранда обнаруживаются в разных молекулярных формах как внутри тромбоцитов (в органеллах), так и на их наружных мембранах. Из собственно тромбоцитарных факторов для свертывания крови наибольшее значение имеет фосфолипидный компонент, или 3-й пластиночный фактор (ПФ-3), представляющий собой организованные в микромембраны липидно-белковые комплексы, на которых, как на матрицах, организуется и ускоряется взаимодействие плазменных факторов свертывания. Второе важное свойство ПФ-3 состоит в том, что, фиксируя на себе активированные факторы IX и X, он защищает их от инактивации наиболее мощным физиологическим антикоагулянтом – антитромбином III и комплексом антитромбин III – гепарин. ПФ-3 включается в процесс свертывания (становится доступным) при активации тромбоцитов, сопряженной с «реакцией освобождения». При некоторых формах патологии тромбоцитов этот механизм нарушается. Сходным с ПФ-3 активирующим действием на свертывание крови обладают мембранные факторы эритроцитов (эритроцитин, эритрофосфатид), активность которых также выявляется в зоне гемостаза. Для исследования свертывающей системы крови используется также тканевый заменитель ПФ-3 – кефалин (вытяжка из ткани мозга, лишенная тромбопластических свойств дополнительным извлечением). Из других тромбоцитарных факторов наиболее важен фактор 4 (ПФ-4) – 2 низкомолекулярных белка, содержащихся в гранулах II типа, с высокой антигепариновой активностью, а также способностью потенцировать агрегацию кровяных пластинок и эритроцитов. Уровень ПФ-4 в плазме является одним из маркеров внутрисосудистой активации тромбоцитарного гемостаза. Заслуживают упоминания также фибринопластический компонент кровяных пластинок, повышающий чувствительность фибриногена к тромбину, фактор ускорения полимеризации фибрин-мономеров и тромбостенин. С его функцией связаны такие важные феномены, как изменение формы кровяных пластинок, образование ложных ножек, «реакция освобождения», трансформация АТФ в АДФ, фиксация тромбоцитов на субэндотелии, укрепление пластиночного тромба и ретракция кровяного сгустка. Активируют процесс свертывания крови и другие вещества, освобождающиеся при активации тромбоцитов, в частности АДФ и адреналин, которые существенно ускоряют переход факторов XII и XI в активированную форму. Тромбоциты оказывают также разнонаправленное влияние на фибринолиз, причем в одних условиях они его ингибируют, а в других, наоборот, активируют. Цельные плазменные сгустки растворяются значительно медленнее в присутствии тромбоцитов, чем без них, но при исследовании растворенных сгустков, полученных из разведенной цельной плазмы, выявляется очень выраженное активирующее влияние тромбоцитов на растворение. Система свертывания крови Механизм гемокоагуляции Основы ферментной теории свертывания крови были заложены еще в XIX в. профессором Юрьевского университета А. А. Шмидтом (1861 г.; 1895 г.) и уточнены П. Моравитцем в 1905 г. Согласно данной теории образование волокон фибрина, составляющих каркас любого свертка крови, связано с ферментным отщеплением от молекул фибриногена небольших фрагментов (фибринопептидов), после чего остающиеся основные части этих молекул (фибрин-мономеры) соединяются друг с другом в длинные цепи «фибринполимера». Фермент крови, обеспечивающий отщепление фибрино-пептидов и превращение фибриногена в фибрин, получил название тромбина. Готового тромбина в плазме нет, но в ней имеется его неактивный предшественник – протромбин (фактор II), который в присутствии ионов кальция и под влиянием «тромбокиназы» превращается в тромбин. Имеется 2 различных механизма активации свертывания крови. Один из них обозначается как «внешний механизм», поскольку запускается поступлением из тканей или из лейкоцитов в плазму тканевого тромбопластического фактора (фактора III), относящегося к липопротеидам. Этот фактор вступает во взаимодействие с фактором VII и при участии ионов кальция быстро образует активатор фактора X, который и является главной составной частью протромбиназы, поскольку трансформирует протромбин (фактор II) в тромбин (IIа). В лабораторных условиях этот путь имитируется протромбиновым тестом Квика: к исследуемой рекальцифицированной плазме добавляется стандартная доза тканевого (мозгового) тромбо-пластина, получается, что процесс искусственно запускается по внешнему механизму. Второй путь активации свертывания назван внутренним, поскольку осуществляется без добавления извне тканевого тромбопластина, за счет внутренних ресурсов плазмы. В искусственных условиях свертывание по внутреннему механизму наблюдается тогда, когда кровь, извлеченная из сосудистого русла, самопроизвольно свертывается в пробирке. Запуск этого внутреннего механизма начинается с активации фактора XII (фактора Хагемана). Эта активация возникает в разных условиях: вследствие контакта крови с поврежденной сосудистой стенкой (коллагеном и другими структурами), с измененными клеточными мембранами, под влиянием некоторых протеаз и адреналина, а вне организма – вследствие контакта крови или плазмы с чужеродной поверхностью – стеклом, иглами, кюветами и др. Этой контактной активации не препятствует удаление из крови ионов кальция, в связи с чем она происходит и в цитратной (или оксалатной) плазме. Однако в этом случае процесс обрывается на активации фактора IX, для которой уже необходим ионизированный кальций. Вслед за фактором XII последовательно активируются факторы XI, IX и VIII. Последние два фактора образуют продукт, который активирует фактор X, что приводит к формированию протромбиназной активности. Вместе с тем сам по себе активированный фактор X обладает слабой протромбиназной активностью, но она усиливается в 1000 раз акселерирующим фактором – фактором V. Точно так же действие фактора IX на фактор X усиливается в несколько тысяч раз фактором VIII – антигемофильным глобулином. Этим обосновывается деление плазменных факторов свертывания на 2 группы: ферментную – факторы XII, XI, IX, VII, X и II и неферментную – факторы I, V и VIII. Фактор X последовательно отщепляет от протромбина два фрагмента, в результате чего образуется тромбин-эстераза, отщепляющая от ?– и ?-цепей фибриногена вначале 2 пептида А, затем – 2 пептида В (всего 4 фибринопептида). Незавершенный фибрин-мономер, от которого отделились лишь пептиды А, обозначается как «дес-А-фибрин», а лишенный пептидов А и В – как «дес-АВ-фибрин». Фибрин-мономеры имеют трехмодулярную структуру, их сборка в полимер проходит этапы формирования димеров, из которых путем дальнейшего продольного и поперечного связывания образуются протофибрилы фибрина. Соединяясь друг с другом, протофибрилы формируют волокна фибрина. Фибринстабилизирующий фактор XIII (плазменная трансглутаминаза) «прошивает» фибрин-полимеры дополнительными перекрестными связями между ?-цепями и тем самым укрепляет фибрин, делает его нерастворимым в мочевине, монохлоруксусной кислоте и других растворителях. Основным активатором фактора XIII является тромбин. В условиях патологии процесс полимеризации фибрина легко нарушается либо вследствие плохой трансформации дес-А-фибрина в дес-АВ-фибрин, либо из-за нарушения сборки димеров и протофибрил. В этих случаях фибрин-мономеры (дес-А-фибрин и дес-АВ-фибрин) соединяются с фибриногеном, образуя средне– и крупномолекулярные (от 450 000 до 2 000 000 и более) растворимые фибрин-мономерные комплексы. Фибриноген в этих комплексах блокируется и утрачивает способность свертывания под влиянием тромбина. Этот феномен, имеющий большое диагностическое значение, в литературе обозначается по-разному – «растворимые фибрин-мономерные комплексы» (РФМК), «фибринемия», «несвертывающийся фибрин», «заблокированный, или тромбинрезистентный, фибриноген», «феномен паракоагуляции». Последнее название связано с тем, что не свертывающиеся тромбином РФМК коагулируют или преципитируют под влиянием ряда неферментных воздействий – при добавлении к плазме спирта (этаноловый тест), сульфата протамина (протамин-сульфатный тест) или при охлаждении (криофибриноген). Биологический смысл и санационное значение образования РФМК заключаются в том, что они способствуют поддержанию жидкого состояния крови при тромбинемии, препятствуют отложению больших масс фибрина в сосудах и уменьшают блокаду зоны микроциркуляции. Вместе с тем доказано, что РФМК значительно легче и быстрее растворяются плазмином, чем коагулировавший фибрин. Таким образом, свертывание крови – многоэтапный каскадный ферментный процесс, в котором последовательно активируются проферменты и действуют силы аутокатализа, функционирующие как сверху вниз, так и по механизму обратной связи. Так, первые малые дозы тромбина, чаще всего образующиеся благодаря включению внешнего механизма свертывания под влиянием тканевого тромбопластина, активируют акселераторы – факторы VIII и V, в результате чего интенсифицируется основный внутренний механизм формирования протромбиназной активности и тромбина. Эти механизмы аутокатализа действуют интенсивно, но кратковременно. Вскоре их сменяют инактивация факторов свертывания и самоторможение системы. Этому способствуют как физиологические антикоагулянты, так и конечные и побочные продукты свертывания, многие с высокой противосвертывающей активностью. Ингибирующим влиянием на свертывание крови и тромбоцитарный гемостаз обладают и продукты фибринолиза. По всем этим причинам свертывание крови затормаживается и не переходит в обычных условиях из локального процесса во всеобщую коагуляцию циркулирующего фибриногена. Также в активации начальных этапов свертывания участвуют компоненты калликреин-кининовой системы. Стимуляторами являются фактор ХIIа и его фрагменты, образующиеся в результате расщепления фактора XII калликреином. Комплекс фактор ХIIа – калликреин – высокомолекулярный кининоген (ВМК) ускоряет активацию не только фактора XI, но и фактора VII, реализуя взаимосвязь между внутренним и внешним механизмами свертывания. Еще более важно активирующее влияние тромбопластина и фактора VII на фактор IX, в результате чего даже малые дозы тканевого тромбопластина запускают процесс свертывания крови не только по внешнему, но и по внутреннему пути, через фактор IX. Установлено также, что фактор ХIIа и его фрагменты через калликреин-кининовую систему, а отчасти и непосредственно активируют ряд других плазменных ферментных систем, в том числе фибринолитическую и систему комплемента. Фактор VIII – многокомпонентная система, состоящая из нескольких субъединиц, участвующих в формировании его коагуляционной активности (VIII : С) и в тромбоцитарно-сосудистом взаимодействии (фактор Виллебранда VIII : FW). Эти субъединицы отличаются разной стабильностью, гетерогенны по генетическому контролю и антигенным свойствам. Противосвертывающие механизмы Противосвертывающие механизмы играют ведущую роль в поддержании жидкого состояния крови и в ограничении процесса тромбообразования. Однако они изучены значительно меньше, чем процесс свертывания крови, в связи с чем вопросы функции и физиологической регуляции антикоагулянтного звена системы гемостаза во многом остаются дискуссионными. Все образующиеся в организме антикоагулянты можно разделить на 2 группы:
Физиологические антикоагулянты, образующиеся независимо от протеолиза. К ним относятся белковые и фосфолипидные ингибиторы начальной фазы свертывания крови, из которых наиболее активен и физиологически важен относящийся к ?2-глобулинам ингибитор фактора XIа. Слабее действует на начальные фазы свертывания липидный антикоагулянт. Из всех предшествующих антикоагулянтов наиболее активен, универсален по действию и важен для поддержания жидкого состояния крови антитромбин III (AT III). Этот белок, содержащийся в плазме в количестве около 0,3–0,42 г/л, или 3,0–4,7 ммоль/л, инактивирует не только тромбин, но и все другие активированные ферментные факторы свертывания: ХIIа, XIa, IXa, Ха. Он же является плазменным кофактором гепарина – без AT III гепарин почти совершенно не оказывает антикоагулянтного действия. Дефицит AT III, наследственный или вторичный (симптоматический), закономерно приводит к развитию тяжелейшего, часто несовместимого с жизнью тромбоэмболического синдрома. Дефицит всех других предшествующих физиологических антикоагулянтов по раздельности либо в совокупности не создает подобных критических ситуаций. Все вышеперечисленные данные закрепили за AT III репутацию главного ингибитора и модулятора системы свертывания крови. Местом синтеза AT III долгое время считали печень, однако исследования последних лет, в том числе выполненные на клеточных культурах иммунологическими методами, показали, что этот антикоагулянт продуцируется сосудистым эндотелием. Все факторы свертывания AT III инактивирует, образуя с ними эквимолярные комплексные соединения. Гепарин, соединяясь с AT III, резко ускоряет это взаимодействие и фиксирует антикоагулянт на поверхности эндотелиальных клеток, чем повышается тромбоустойчивость внутренней стенки сосудов. Альфа2-макроглобулин является слабым ингибитором тромбина, действие которого становится ощутимым лишь при депрессии AT III. На долю этого антикоагулянта приходится, по разным авторам, от 4 до 21% антитромбиновой активности дефибринированной плазмы. Несколько больше роль ?2-макроглобулина в связывании плазмина, но и в этом случае его действие становится ощутимым после удаления быстродействующего антиплазмина. В отличие от AT III он из всех активированных факторов свертывания взаимодействует только с тромбином. Наследственный дефицит ?2-макроглобулина не сопровождается ни сколько-нибудь заметной тромбогенностью, ни существенными сдвигами в свертывающей системе крови, что говорит о его весьма ограниченном значении в регуляции гемостатического потенциала крови. Более выражено ингибирующее действие на тромбин и другие активированные факторы свертывания (IXa, XIa и ХПа) ?1-антитрипсина и ингибитора 1 компонента комплемента. Однако и при их дефиците не наблюдается значительных нарушений гемостаза, что, очевидно, связано с одинаково выраженным ослаблением инактивации как свертывания крови, так и фибринолиза, вследствие чего сохраняется динамическое равновесие между этими системами. Антикоагулянты, образующиеся в процессе свертывания крови и фибринолиза. Многие прокоагулянты и их метаболиты в процессе свертывания крови и фибринолиза приобретают антикоагулянтные свойства. Так, фибрин адсорбирует и инактивирует образующийся при свертывании тромбин, вследствие чего фибрин обозначается как антитромбин I. Эта инактивация настолько велика, что в сыворотке, как известно, остаются ничтожно малые количества тромбина. Имеются указания, что фибринопептиды, отщепляемые от фибриногена тромбином, также обладают антикоагулянтным действием. Самоторможение наблюдается и на других этапах свертывания. Так, тромбин действует ферментативно на протромбин, отщепляя от него ингибитор фактора Ха; фактор Va после участия в свертывании начинает тормозить превращение протромбина в тромбин, а фактор ХIа после взаимодействия с фактором XII начинает тормозить его дальнейшую активацию. Мощные антикоагулянты, обладающие антитромбиновым и антиполимеразным действием, образуются в процессе фибринолиза. Все вышеперечисленные данные в очередной раз свидетельствуют о том, что в свертывающей системе крови на всех этапах каскада действуют силы самоограничения процесса, одни и те же факторы могут выступать вначале как коагулянты, а затем – как антикоагулянты. Схема 2 Система свертывания крови 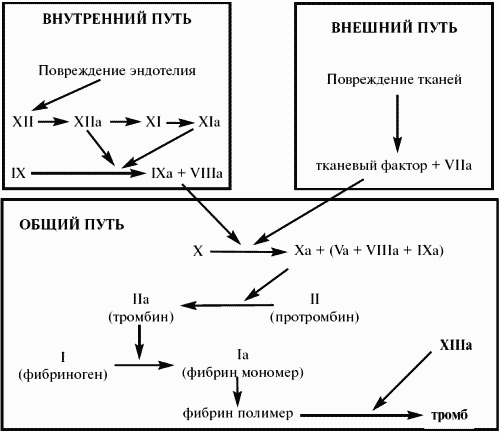 Фибринолитическая (плазминовая) система Ферментная система, обеспечивающая растворение фибрина в кровяном русле, получила название фибринолитической, или плазминовой, системы. Это растворение осуществляется основным компонентом указанной системы – фибринолизином (или плазмином), который в плазме содержится в виде профермента (плазминогена) в концентрации около 20,6 + 3,6 мг%. Как в плазме, так и в тканях плазминоген содержится в виде двух или более молекулярных форм, отличающихся друг от друга способами выделения, особенностями активации и инактивации. Каждая из двух основных форм состоит из нескольких молекулярных подформ: 1) нативный плазминоген с NH2-терминальной глютаминовой кислотой – «глю-плазминоген»; 2) подвергшийся ограниченному протеолизу плазминоген с NH2-терминальным лизином, валином или метионином – «лиз-плазминоген». Лиз-плазминоген в 10–20 раз быстрее трансформируется активаторами в плазмин, имеет значительно более выраженное, чем глю-плазминоген, сродство к фибрину и значительно быстрее последнего метаболизируется – его Т1/2 в циркуляции около 0,8 суток, а глю-плазминогена – 1,24 ± 0,29 суток. По механизму протеолитического действия плазмин наиболее близок к трипсину. После активации плазминоген быстро исчезает из кровотока – блокируется антиплазминами и удаляется. Вслед за введением больших доз стрептокиназы или урокиназы уровень плазминогена в крови снижается до нуля, но затем в течение 12–28 ч восстанавливается, если прекращена его дальнейшая активация. Эта способность активаторов фибринолиза быстро истощать запасы плазминогена в крови и на время оставлять больного без ферментативного фибринолиза важна для клиники и должна учитываться при лечении тромбозов и синдрома диссеминированного внутрисосудистого свертывания крови. Существующие в организме механизмы активации плазминогена весьма разнообразны, но, подобно механизмам свертывания крови, они также могут быть подразделены на две основные группы – с внутренней и внешней активацией. Ведущий внутренний механизм запускается теми же факторами, какие инициируют свертывание крови, а именно фактором ХIIа, который, взаимодействуя с прекалликреином и высокомолекулярным кининогеном плазмы (ВМК), активирует плазминоген. Такой путь фибринолиза – базисный, обеспечивающий активацию плазминовой системы не вслед за свертыванием крови, а одновременно с ним. Он функционирует по «замкнутому циклу», поскольку образующиеся первые порции калликреина и плазмина вызывают протеолиз фактора XII, отцепляя фрагменты, под действием которых нарастает изменение прекалликреина в калликреин. Такая интенсивная самоактивация приводит к тому, что ХIIа-калликреин-зависимый фибринолиз при интенсивном внутрисосудистом свертывании крови истощается быстро, раньше других механизмов фибринолиза. Лимитирующими факторами являются в первую очередь ВМК и прекалликреин. Их плазменный резерв быстро истощается, тогда как уровень плазминогена остается в крови еще достаточно высоким. В таких условиях ХIIа-зависимый фибринолиз уже не функционирует, но поддается другим (не калликреиновым) способам активации – стрептокиназой и урокиназой. Лишь вслед за этим возможно истощение запасов плазминогена, что делает неэффективным любые способы активации плазминовой системы. Определенное участие в активации внутреннего механизма фибринолиза принимает, по-видимому, и фактор Виллебранда. В частности, на образцах плазмы с дефицитом ВМК показано, что фактор Виллебранда в 2–3 раза усиливает превращение прекалликреина в калликреин под влиянием фрагментов фактора XII. В присутствии ВМК, обладающего значительно более мощным влиянием на активацию прекалликреина, это действие фактора Виллебранда становится малоощутимым. Заслуживает внимания то обстоятельство, что если в свертывании крови компонентам калликреин-кининовой системы отводится в определенной мере вспомогательная функция, то в гуморальном механизме фибринолиза это один из ведущих механизмов. Возможно, именно поэтому при генетически обусловленном дефиците плазменного прекалликреина (дефект Флетчера) или ВМК (дефект Фитцжеральда – Вильсона) у больных нет кровоточивости и вместе с тем прослеживается наклонность к тромбозам. Важнейшими стимуляторами внешнего механизма фибринолиза являются белковые активаторы плазминогена, синтезируемые в сосудистой стенке. Эти активаторы подразделяются на высокомолекулярные и низкомолекулярные фракции, обнаруживают высокое сродство к фибрину. Физиологическая регуляция синтеза и выделения в кровь сосудистых активаторов изучена недостаточно. Тем не менее известно, что их интенсивный выброс происходит при нарушении проходимости сосудов, в том числе и при пережатии сосуда манжетой, а также при физических нагрузках, под влиянием вазоактивных веществ. Определение эуглобулинового лизиса до и после пережатия сосуда (манжеточная проба) используется для оценки резерва сосудистых активаторов плазминогена и функциональной полноценности механизмов их либерации. Депрессия данных механизмов характерна для ряда тромбофилических состояний. Стероидные гормоны анаболического действия повышают синтез в эндотелии активаторов фибринолиза, с чем отчасти связывается их благоприятное влияние на течение флеботромботической болезни. Мощные активаторы плазминогена содержатся также в клетках крови – эритроцитах, тромбоцитах и особенно лейкоцитах. При внутрисосудистом свертывании крови, тромбообразовании, воздействии эндотоксином, активации системы комплемента, гемолизе эти активаторы освобождаются из клеток в «плазматическую атмосферу» и активируют плазминоген. Более того, установлено, что гранулоциты секретируют не только активатор плазминогена, но и внутриклеточные протеазы (цитокиназы), которые самостоятельно, без участия плазмина, переваривают фибрин. При этом образуются иные продукты расщепления фибрина, чем при его плазминовом расщеплении. Следовательно, лейкоциты обеспечивают функционирование самостоятельного (неплазминового) механизма растворения фибрина. Этот альтернативный механизм играет важную роль в ограничении размеров тромбов и в деблокировании микроциркуляторного русла при диссеминированном внутрисосудистом свертывании крови. Разнообразные активаторы плазминогена (цитокиназы) содержатся и в других тканях и клетках, особенно в эпителиальной, мышечной и мезенхимальной, а также в секретах и экскретах – моче, молоке, желчи, слюне. Некоторые из них поступают в определенных количествах в кровь, участвуя в активации плазминогена. В частности, таким свойством обладает урокиназа – активатор фибринолиза, синтезируемый в почечном эпителии и выделяющийся с мочой. В кровь поступает небольшое количество урокиназы, ответственное приблизительно за 10–15% общей плазминоген-активаторной функции. В настоящее время установлено, что большинство тканевых активаторов плазминоген идентично сосудистому, эндотелиальному. Фибринолиз ингибируется рядом антиактиваторов и антиплазминов, из которых наиболее важен недавно открытый быстродействующий антиплазмин, относящийся к ?2-глобулинам (молекулярная масса 65 000–70 000) и содержащийся в плазме в количестве 70 мг/л. Этого количества достаточно, чтобы нейтрализовать более 2/3 всего плазмина, образующегося при максимальной активации плазминогена. Однако плазмин, связанный с фибрином, хуже комплексируется с антиплазмином, чем при циркуляции в свободном состоянии. Антиплазмин ослабляет процесс связывания плазминогена с фибрином. Присутствие в плазме циркулирующих комплексов плазмин – антиплазмин, как и комплексов тромбин-ATIII, служит признаком интенсивного внутрисосудистого свертывания крови и активации фибринолиза. Выявление этих комплексов облегчается тем, что в них появляются новые антигенные свойства (так называемые неоантигены). Быстродействующий ?2-антиплазмин обладает также антиактиваторным действием, но он не идентичен другому антиактиватору, описанному Hedner (1973, 1977 гг.). Из других ингибиторов фибринолиза, обладающих значительно более слабым действием, заслуживают упоминания ?2-макроглобулин и ингибитор С1-эстеразы. Последний ингибирует фактор ХIIа, калликреин и отчасти плазмин, специфически блокирует внутренний (ХIIа-зависимый) фибринолиз. Вместе с тем имеются данные о том, что ?2-макроглобулин не столько препятствует фибринолизу, сколько защищает плазмин от других, более мощных, ингибиторов. В частности, комплекс макроглобулин-плазмин защищен от быстродействующего ?2-антиплазмина, благодаря чему при активации плазминовой системы идет лизис не только фибрина и РФМК, но в небольшой степени и фибриногена, хотя в плазме имеется избыток ?2-антиплазмина. Плазминовая система специфически адаптирована к лизису фибрина и растворимых фибрин-мономерных комплексов (РФМК), хотя при ее значительной активации расщеплению подвергаются и другие белки (в том числе факторы свертывания V и VIII). Механизм преимущественной активации фибринолиза в тромбах и сгустках, резко выраженного преобладания фибринолиза над фибриногенолизом пока не может считаться окончательно выясненным. Твердо доказана лишь способность частично активированного плазминогена (лиз-плазминогена) связываться с фибрином. Установлено также, что растворение идет тем быстрее, чем выше локальная концентрация в сгустках плазминогена. Особенно важно, что сосудистый активатор плазминогена также концентрируется на фибрине. Наконец, установлено, что ?2-антиплазмин намного слабее инактивирует связанный с фибрином плазмин, тогда как циркулирующий «свободный» плазмин образует с этим мощным ингибитором плохо диссоциирующие комплексы. Схема 3 Фибринолитическая система 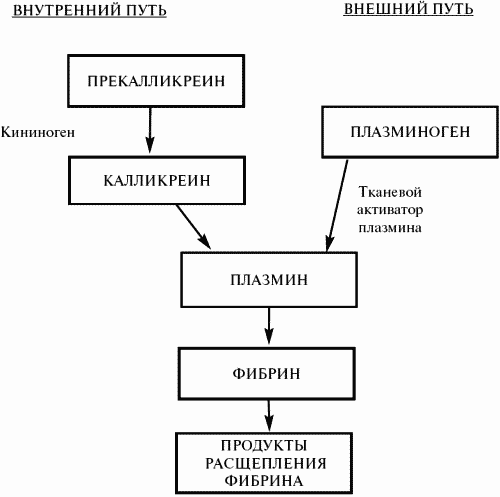 |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||