|
||||
|
|
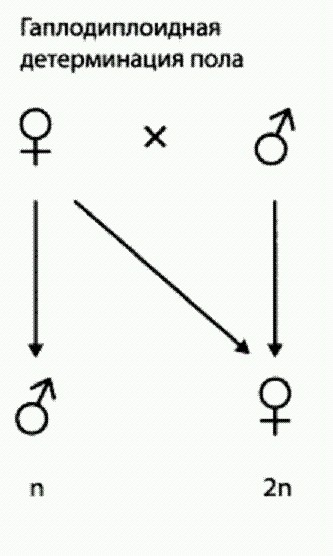
ГЛАВА 5. ЭВОЛЮЦИЯ АЛЬТРУИЗМАРодственный отборКооперация и альтруизм — краеугольные камни социального поведения Homo sapiens. Вряд ли нужно объяснять, что без кооперации (сотрудничества, взаимовыгодного поведения) устойчивое существование социума у приматов невозможно. Очевидно и то, что наши предки эволюционировали именно как социальные, общественные животные. Социальная жизнь часто предполагает и некоторую долю альтруизма, жертвенности. Это не абсолютное правило, но в эволюции гоминид оно, по всей видимости, соблюдалось неукоснительно. Уже 1,7 млн лет назад ранние Homo заботились о беззубых стариках (см. главу "Очеловечивание" в кн. 1). А ведь это альтруизм — кормить беспомощного старика, вместо того чтобы съесть эту пищу самим или отдать своим детям. Поэтому мы не поймем эволюцию человека, пока не разберемся в эволюционных механизмах, ведущих к развитию кооперативного и альтруистического поведения. Это тем более важно, что до сих пор еще приходится иногда слышать от далеких от биологии людей, что эволюция якобы не может объяснить альтруизм. На самом деле эволюция превосходно его объясняет. Каким образом — в этом мы сейчас попробуем разобраться. Изучение эволюции альтруизма и кооперации — это одно из тех направлений, двигаясь по которым биология — естественная наука — вторгается на территорию, где до сих пор безраздельно хозяйничали философы, теологи и гуманитарии. Неудивительно, что вокруг эволюционной этики (так называют это научное направление) кипят страсти. Об этих страстях мы говорить не будем, потому что они кипят за пределами естественных наук. Пусть себе кипят. Нас, биологов, интересует другое. Нас интересует, почему, с одной стороны, большинство живых существ ведут себя эгоистично, но при этом немало есть и таких, кто совершает альтруистические поступки, то есть жертвует собой ради других. Альтруизм у дрожжей? Да как вы смеете! Часто приходится слышать, что такие понятия, как альтруизм (а еще мышление, желание, планирование, понимание, любовь и многие-многие другие), применимы только к человеку и больше ни к кому. По-моему, это просто дело привычки и общественной договоренности. Главное — чтобы было понятно, о чем речь. Слова для людей, а не люди для слов. Было время, даже слово "поведение" кое-кто считал допустимым применять только к людям. Ну как же: ведь только люди могут вести себя осознанно, осмысленно, то есть по-настоящему. А остальные живые существа себя не осознают, личности у них нет, и поэтому никак "себя вести" они не могут. Сейчас такие рассуждения выглядят историческим курьезом — все давно привыкли, что у нечеловеческих животных тоже есть поведение. Иначе можно договориться до того, что только люди могут есть: ведь мы едим осознанно, а все остальные — неосознанно. Употребление слова "альтруизм" по отношению к нечеловеческим живым существам имеет в биологической литературе давнюю историю. Но все-таки многие еще не успели к этому привыкнуть. Поэтому нужно немного разобраться с определениями, тем более что общепринятый смысл понятия "альтруизм" в применении к людям (в этике) и другим живым существам (в биологии) не совсем одинаков. АЛЬТРУИЗМ (ЛАТ. ALTER — ДРУГОЙ) В ЭТИКЕ — нравственный принцип, предписывающий бескорыстные действия, направленные на благо других людей; способность приносить свою выгоду в жертву ради общего блага. АЛЬТРУИЗМ В БИОЛОГИИ: поведение, ведущее к повышению приспособленности (репродуктивного успеха) других особей в ущерб своим собственным шансам на успешное размножение. В этих определениях много общего. В обоих случаях имеется в виду принесение индивидом в жертву своих интересов ("выгоды") ради кого-то другого. Однако интересы могут быть разные. В случае с человеком подразумевается, что у индивида есть какие-то цели, которые он в той или иной степени осознает (и может быть, даже сам себе их поставил, хотя что это значит — "сам поставил" — вопрос очень каверзный; см. раздел "Свободная воля" в главе "В поисках душевной грани"). Причем здесь имеются в виду не любые цели, а корыстные, эгоистические, связанные с личной выгодой (благами, удовольствиями). И наш индивид вдруг берет и отказывается от осуществления своей корыстной цели — жертвует своей "выгодой" ради кого-то другого. Вот вам и альтруизм. Это определение (как и любое другое) можно при желании свести к абсурду и разрушить всю его слабенькую логику, если начать детализировать и докапываться: а что такое благо, а что такое выгода, а что такое удовольствие, а не получает ли человек удовольствие, совершая добрый поступок, а если получает, то это уже не альтруизм, и т.д. Центральную роль в принятии решений у нас все-таки играют подкорковые структуры с их контурами боли и награды, кнута и пряника. Кора больших полушарий выполняет сложные калькуляции, строит модели, прогнозирует последствия возможных поступков, сравнивает их с последствиями альтернативных вариантов поведения. Но эмоциональная оценка результатов всех этих вычислений зависит от подкорковых структур. Это они в конечном счете определяют, что делать следует, а что нет. Какой из вариантов даст в итоге больше пряников-эндорфинов. Хвостатое ядро капризно виляет хвостиком: хочу — не хочу, буду — не буду. Если вам хочется совершить добрый поступок (и получить на свою голову все его предполагаемые последствия) сильнее, чем хочется его не совершать (тоже со всеми вытекающими), то вы его совершите. Если мысленная модель планируемого доброго поступка активирует нейронную сеть "кнута" сильнее, чем нейронную сеть "пряника", то вы его не совершите. Так что при желании можно привести аргументы в поддержку тезиса, что альтруизма вообще не бывает: что хотим, то и делаем. Можно порассуждать о том, что моральные нормы, законы, системы запретов и табу как раз для того и выработаны обществом, чтобы наши хвостатые ядра чаще склоняли хвост в сторону поступков, полезных окружающим. Но лучше не будем погружаться в эти демагогические пучины. Остановимся на том, что большинству людей интуитивно понятно, что такое "эгоистический интерес", когда дело касается человека. Перейдем к биологическому определению. Какие цели, какие интересы могут быть у дрожжевой клетки? Строго говоря, никаких интересов у нее нет и целей тоже. Но нам удобно думать, что у нее есть одна, совершенно определенная цель: размножиться, растиражировать свои гены в следующих поколениях. Нам удобно приписать ей такую цель, потому что если приглядеться внимательно к любому живому существу, то мы увидим, что оно замечательно приспособлено для выполнения именно этого действия — размножения, тиражирования своих генов. Так получается само собой, потому что так работает естественный отбор. Если случайная мутация повышает эффективность тиражирования данного фрагмента ДНК, то (как можно догадаться) данный фрагмент ДНК будет тиражироваться эффективнее. Причем, что важно, тиражироваться он будет вместе с этой самой мутацией, которая повысила эффективность его размножения. Число копий гена с такой мутацией будет расти. Если другая мутация снизит эффективность размножения своего кусочка ДНК, этот кусочек — вместе с этой мутацией — будет размножаться хуже, и число его копий в этом мире будет снижаться. В результате мир неуклонно наполняется фрагментами ДНК, все более эффективно себя тиражирующими. Ни у кого нет никаких целей. Однако выглядит все так, как будто у фрагментов ДНК (генов) есть цель: размножаться как можно эффективнее. На самом деле они просто автоматически, в силу слепых законов природы, накапливают изменения, повышающие эффективность их размножения. Но нам — в силу некоторых особенностей нашей психики — удобно рассматривать этот процесс как целенаправленный. В действительности он просто направленный, без "целе-". Естественный отбор придает ему направленность в сторону роста эффективности размножения генов. А цель ему приписываем мы. Нам так удобно, потому что мы так устроены: мы любим приписывать цели направленным процессам, результаты которых можно в какой-то мере предвидеть. Или объектам, от которых можно ожидать более-менее определенных действий. Философ Дэниел Деннетт объяснил, почему это нам удобно, в книге The Intentional Stance ("Интенциональная установка"). Это ценная психическая адаптация, удобное "срезание угла", кратчайший путь к практичному выводу, лихой прыжок через длинную цепочку "лишних" логических рассуждений — нейронных калькуляций. Если в джунглях на нас бросается хищник, глупо размышлять о природе его поведенческих реакций, о его рецепторах, лимбической системе и гормональном статусе. "Он хочет меня сожрать!" — вот как должен осмыслить поведение хищника жизнеспособный, адаптированный к реалиям этого мира двуногий примат. В некоторых языках даже будущее время формируется на основе глагола "хотеть": говорят не "поезд придет через пять минут", а "поезд хочет прийти через пять минут". Нам удобно использовать интенциональную установку, потому что у нас великолепно развита "теория ума". У нас есть хорошая мысленная модель себя, и нам легко судить о внешних объектах по себе. А мы-то сами постоянно занимаемся планированием и ставим себе цели (по крайней мере так нам кажется). Раз нам это удобно, то давайте так и поступим. Будем считать понарошку, что у генов и организмов есть цель — как можно эффективнее размножиться. Пусть это будет их "корыстным интересом". А когда организм вдруг начинает вести себя в ущерб своему корыстному интересу, да притом еще так, что это идет на пользу интересам другого организма, мы будем называть такое поведение альтруистическим. Вот и ладно, вот и договорились. Итак, перед нами стоят два основных вопроса. С одной стороны, ясно, что многие жизненные задачи легче решать совместными усилиями, чем в одиночку. Почему же тогда биосфера так и не превратилась в царство всеобщей дружбы и взаимопомощи? Это первый вопрос. Второй вопрос противоположен первому. Как вообще может в ходе эволюции возникнуть альтруизм, если движущей силой эволюции является естественный отбор — процесс, как представляется на первый взгляд, абсолютно эгоистический? Все дело в том, что этот "первый взгляд" — неправильный. Ошибка здесь в смешении уровней, на которых мы рассматриваем эволюцию. Эволюцию можно рассматривать на разных уровнях: генов, индивидов, групп, популяций, экосистем, всей биосферы. На каждом уровне свои закономерности и правила. На уровне генов в основе эволюции лежит конкуренция разных вариантов (аллелей) одного и того же гена за доминирование в генофонде популяции. На генном уровне никакого альтруизма нет и быть не может. Ген всегда эгоистичен. Если появится "добрый" аллель, который в ущерб себе позволит размножаться другому аллелю, то этот альтруистический аллель будет вытеснен из генофонда и просто исчезнет. Но если мы переведем взгляд с уровня генов на уровень организмов, то картина будет уже другой. Потому что интересы гена не всегда совпадают с интересами организма. Ген, или, точнее, аллель, — это не единичный объект, он присутствует в генофонде в виде множества одинаковых копий. "Интерес" у всех этих копий один и тот же. Ведь они — просто молекулы, и они абсолютно идентичны. И им, и нам, и естественному отбору совершенно все равно, какая именно из одинаковых молекул размножится, а какая нет. Важен только суммарный итог: сколько копий аллеля было и сколько их стало. Организм, напротив, — это единичный объект, и в его геноме могут присутствовать, говоря упрощенно, только одна или две копии интересующего нас аллеля. Иногда эгоистичному гену выгодно пожертвовать одной- двумя своими копиями для того, чтобы обеспечить преимущество остальным своим копиям, которые заключены в других организмах. К этой мысли биологи стали подходить уже в 30-е годы прошлого века. Важный вклад в понимание эволюции альтруизма внесли Рональд Фишер (Гениальный биолог-теоретик, один из создателей генетической теории эволюции и современной статистики. У него были свои недостатки: рассказывают, что он заставлял жену рожать ребенка за ребенком, потому что считал, что у него шикарные гены, которые необходимо растиражировать. Фишер был поклонником евгеники), Джон Холдейн (Тоже великий биолог. Прославился своими афоризмами. Однажды в ответ на заявление некой дамы, которая не могла поверить, что "из одноклеточного организма даже за миллиарды лет может развиться такое сложнейшее существо, как человек", Холдейн сказал: "Мадам, вы сами это проделали. И у вас ушло на это всего девять месяцев". В другой раз на вопрос одного богослова, какая черта личности Создателя наиболее ярко проявилась в его Творении, Холдейн ответил: "Необычайная любовь к жукам" (жуки - самая разнообразная группа живых существ, их известно более 300 000 видов)) и Уильям Гамильтон (Тоже гений. Возможно, величайший биолог-теоретик XX века. Разработал и математически обосновал теорию родственного отбора, эволюционную теорию старения и много других замечательных идей). Теория, которую они построили, называется теорией родственного отбора. Суть ее образно выразил Холдейн, который однажды сказал: "Я бы отдал жизнь за двух братьев или восьмерых кузенов". Что он имел при этом в виду, можно понять из формулы, которая вошла в науку под названием "правило Гамильтона". Вот эта формула. "Ген альтруизма" (точнее, аллель, способствующий альтруистическому поведению) будет поддержан отбором и распространится в популяции, если RB > С, где R — степень генетического родства жертвователя и "принимающего жертву" (на самом деле родство важно не само по себе, а только как фактор, определяющий вероятность того, что у "принимающего" имеется тот же самый аллель альтруизма, что и у жертвователя); В — репродуктивное преимущество, полученное адресатом альтруистического акта; С — репродуктивный ущерб, нанесенный "жертвователем" самому себе. Репродуктивный выигрыш или ущерб можно измерять, например, числом оставленных или не оставленных потомков. С учетом того, что от акта альтруизма может выиграть не одна, а много особей, формулу можно модифицировать следующим образом: NRB > С, где N — число принимающих жертву. Обратите внимание, что правило Гамильтона не вводит никаких дополнительных сущностей, не требует специальных допущений и даже не нуждается в экспериментальной проверке. Оно чисто логически выводится из определений величин R, В, С и N — точно так же, как геометрические теоремы выводятся из аксиом. Если NRB > С, "аллель альтруизма" будет совершенно автоматически увеличивать свою частоту в генофонде популяции. Посмотрим, как это работает, что называется, на пальцах. Допустим, наш аллель альтруизма заставляет своего носителя пожертвовать жизнью, если эта жертва спасает жизнь трех его родных братьев (с двумя Холдейн погорячился: два брата — это шило на мыло; почему, будет объяснено ниже). Допустим, это действие совершается в молодом возрасте, когда никто из участников еще не имеет детей, и допустим, что в среднем каждый самец в этой популяции оставляет троих детей. Подсчитаем значения переменных. Число адресатов (N) равно трем (три брата). Родство (R) равно 0,5. Жертвователь получил свой аллель альтруизма от одного из родителей. С каждым из братьев он имеет в среднем 50% общих генов. В данном случае "общих" означает "идентичных по происхождению", то есть представляющих собой точные копии одного и того же родительского гена. Поэтому вероятность того, что данный брат имеет копию того же самого аллеля альтруизма, равна в среднем 0,5. Репродуктивный выигрыш (В) равен трем. Брат оставит троих потомков, если выживет, и ни одного, если погибнет. Репродуктивный ущерб (С) тоже равен трем. Совершив альтруистический акт, жертвователь погибает и не оставляет троих детей, которых он оставил бы, не совершив самопожертвования. Подставив эти числа в неравенство, получаем выражение: NRB = 4,5 > 3. Неравенство истинно, значит, аллель альтруизма при данных условиях должен распространяться. Проверим, так ли это. Если жертва будет принесена, спасенные братья оставят по три потомка. Каждый брат имеет аллель альтруизма с вероятностью 0,5. Каждому из их детей этот аллель, если он у них есть, достанется тоже с вероятностью 0,5. Всего, таким образом, получится девять потомков, каждый из которых имеет аллель альтруизма с вероятностью 0,25. В среднем в следующее поколение перейдет 9 х 0,25 = 2,25 копии аллеля альтруизма. Если жертва не будет принесена, в следующее поколение перейдет в среднем С х 0,5 = 1,5 копий аллеля. Мораль: аллелю выгодно, чтобы жертвователь совершил альтруистический акт. Благодаря этому акту аллель передаст в следующее поколение не 1,5, а 2,25 своей копии. Следовательно, аллель альтруизма с течением времени будет наращивать свою частоту, вытесняя из генофонда конкурирующий "аллель эгоизма". Подставив двух братьев вместо трех, получим равенство: 1,5 = 1,5. С точки зрения самого аллеля никакого альтруизма тут нет, один сплошной эгоизм. Аллель заставляет своих носителей — то есть организмы — жертвовать собой, но тем самым аллель блюдет свои корыстные интересы. Он жертвует небольшим числом своих копий, чтобы дать преимущество большему числу точно таких же своих копий. Естественный отбор — это автоматическое взвешивание суммы выигрышей и проигрышей для аллеля — для всех его копий вместе, и если выигрыши перевешивают, аллель распространяется. Вот, собственно и вся теория. Глупая чайка Правило Гамильтона обладает замечательной объясняющей и предсказательной силой. Например, оно помогает объяснить типичную сцену, которую можно наблюдать на берегах водоемов (я ее часто наблюдаю летом на Белом море). Поймал рыбак рыбку и начинает ее на берегу потрошить, бросая потроха в воду. Это замечает чайка, она прилетает и начинает хватать потроха из воды. Но она делает это не молча, а сначала издает несколько громких призывных криков. На эти крики быстро слетается еще десятка два чаек, которые тут же набрасываются на первую чайку и начинают отнимать у нее добычу. Та не отдает, отбивается, разыгрывается целый спектакль с вырыванием друг у друга из клюва рыбьих потрохов. Странное поведение! С одной стороны, почему бы чайке не есть молча? Зачем она позвала других, создав тем самым себе проблемы? Второй вопрос: если уж она их позвала, то почему тогда не хочет поделиться, а дерется и не отдает? В этой сцене, как и во многих других ситуациях в живой природе, мы видим причудливое сочетание альтруистического и эгоистического поведения. Призывный пищевой крик чайки — типичный пример альтруизма. От этого крика чайка не получает никакой выгоды. Выигрыш достается другим чайкам: они получают шанс пообедать. Вторая часть сцены — драка. Здесь уже, конечно, мы видим лишь чистый эгоизм со стороны всех участников. Разгадка — в правиле Гамильтона. Чайки на Белом море питаются в основном стайными рыбами, например селедкой. Если чайка заметила одну рыбку, то, скорее всего, рядом есть много других: на всех хватит. Это значит, что величина С — цена альтруистического акта — будет в среднем низкой. Величина В — выигрыш тех, кто прилетит на крик, — будет довольно большой: они пообедают. Поскольку рыба стайная, следующую стаю, возможно, придется долго ждать. Величина R (родство) тоже, скорее всего, будет высокой, потому что чайки гнездятся колониями, часто возвращаются на одно и то же место после зимовки, и поэтому, скорее всего, рядом с этой чайкой гнездятся ее родственники — родители, дети, братья и племянники. Величина N — число чаек, которые услышат, прилетят и пообедают, тоже довольно высоко. Неравенство NRB > С выполняется. Вот и закрепились в популяции чаек мутации, способствующие "пищевому крику". А почему чайка не делится своей добычей, не отдает то, что уже схватила? Потому что в этом случае величина С больше: чайка остается без обеда. Величина N, напротив, оказывается меньше. Отдав свою добычу другой чайке, она накормит одну, а не целую стаю. Неравенство не выполняется, и мутации, склоняющие чайку поделиться добычей, не закрепляются, отсеиваются отбором. Конечно, выгоднее всего для чайки (точнее, для ее генов) было бы научиться различать ситуацию, когда пищи много и хватит на всех, и когда пищи мало. В первом случае выгодно кричать, а во втором помалкивать. Но для таких калькуляций нужны мозги. А мозг, как мы знаем, дорогой орган. Отбор, как правило, пытается сэкономить на мозгах. К тому же мозги тяжелые. Чайкам надо летать, а не решать алгебраические задачи. Поэтому птица и не может сообразить, когда ей выгодно звать товарок, а когда нет, и ее поведение оказывается нелогичным. Не всегда, а только при недостатке рыбок. Эволюция альтруизма особенно далеко зашла у перепончатокрылых насекомых: муравьев, пчел, ос, шмелей. У общественных перепончатокрылых большинство самок отказываются от собственного размножения, чтобы выкармливать сестер. Это высшее проявление альтруизма. Таких животных называют эусоциальными, то есть "истинно общественными". Но почему именно перепончатокрылые? Гамильтон предположил, что дело тут в особенностях наследования пола. У перепончатокрылых самки имеют двойной набор хромосом, а самцы — одинарный. Из-за этого складывается парадоксальная ситуация: сестры оказываются более близкими родственницами, чем мать и дочь. У большинства животных сестры имеют 50% общих (идентичных по происхождению) генов. Величина R в формуле Гамильтона равна 1/2. У перепончатокрылых сестры имеют 75% общих генов (R = 3/4), потому что каждая сестра получает от отца не половину его хромосом, а весь геном полностью. Мать и дочь у перепончатокрылых имеют, как и у других животных, лишь 50% общих генов. Вот и получается, что самкам перепончатокрылых при прочих равных выгоднее выращивать сестер, чем дочерей.
Механизм наследования пола у перепончатокрылых. Самка диплоидная, то есть имеет двойной набор хромосом (2n). Она может отложить неоплодотворенное яйцо с одинарным набором хромосом (п), из которого выведется гаплоидный самец. Если же яйцо будет оплодотворено, то его хромосомный набор будет двойным, и из него выведется самка. Самка получает половину хромосом от матери, половину — от отца. Самец получает от матери половину ее хромосом, а отца у него нет. Такой механизм наследования пола называется гаплодиплоидным. В действительности все несколько сложнее. Кроме сестер есть еще и братья-трутни, которые имеют со своими сестрами лишь 25% общих генов (если смотреть со стороны сестры) или 50% (с точки зрения брата). Однако рабочие самки выращивают и братьев тоже (хоть и недолюбливают их). Мы не будем вдаваться в эту довольно сложную теоретическую область, тем более что приматы, которые нас интересуют, не являются гаплодиплоидами. Но у общественных перепончатокрылых есть (или было в эволюционном прошлом) еще одно важное свойство, резко повышающее вероятность развития альтруизма под действием родственного отбора. Это свойство — моногамия. Потомки моногамных диплоидных родителей имеют в среднем по 50% общих генов (R = 0,5). У потомков самки, спаривающейся со многими самцами, средняя величина R стремится к 0,25 (если самцов достаточно много). Для родственного отбора это очень серьезная разница. При R = 0,5 достаточно любого пустяка, чтобы склонить чашу весов в сторону предпочтения сестер и братьев. При R = 0,25 свои дети однозначно дороже. Очень важно, что моногамия свойственна термитам — второму отряду насекомых, в котором эусоциальность получила широкое распространение, причем без всякой гаплодиплоидности. У термитов работают не только самки, но и самцы (они диплоидные, как и их сестры). Как мы помним, моногамия, вероятно, была свойственна древним гоминидам. Это могло стать мощным стимулом для развития под действием родственного отбора братской (и сестринской) взаимовыручки, внутрисемейной кооперации и альтруизма. И еще, конечно, отцовской любви, а заодно и преданности детей обоим родителям, а не только матери. Возможно, всю эту гамму альтруистических чувств родственный отбор смог поддержать у наших предков именно потому, что они были — хотя бы отчасти — моногамными. ОбманщикиКроме родственного отбора существуют и другие механизмы и факторы, помогающие или, наоборот, препятствующие эволюции альтруизма. Главной помехой является проблема так называемых "обманщиков", проблема социального паразитизма. Вспомним, например, социальную жизнь бактерии Pseudomonas fluorescens, о которой говорилось в книге "Рождение сложности". Эта бактерия — удобный объект для изучения эволюции в пробирке. В жидкой среде бактерии Pseudomonas развиваются сначала как одиночные клетки и постепенно занимают всю толщу бульона. Когда в среде становится мало кислорода, получают преимущество бактерии-мутанты, которые выделяют вещества, способствующие склеиванию клеток. Такие бактерии после деления не могут "отклеиться" друг от друга. Фокус тут в том. что одиночные клетки плавают в толще бульона, а склеившиеся всплывают на поверхность, где кислорода гораздо больше, Производство клея — дело дорогостоящее, однако общая награда (кислород) с лихвой покрывает расходы. Возникновение таких колоний — само по себе большое достижение. Но до настоящей социальности, а тем более до настоящего многоклеточного организма тут еще очень далеко. Эти колонии недолговечны, потому что естественный отбор в такой колонии благоприятствует размножению клеток-"обманщиков", то есть мутантов, которые перестают производить клей, однако продолжают пользоваться преимуществами жизни в группе. В этой системе нет никаких механизмов, которые препятствовали бы такому жульничеству. Безнаказанность ведет к быстрому размножению обманщиков, и колония разрушается. Дальнейшее развитие кооперации в такой системе оказывается невозможным из-за социального паразитизма. В этом и состоит главное препятствие на пути эволюции кооперации и альтруизма. Таково общее правило: как только начинает зарождаться кооперация, тут же появляются всевозможные обманщики, нахлебники и паразиты, которые могут лишить кооперацию всякого смысла. Чтобы социальная система могла развиваться дальше, ей необходимо выработать механизм борьбы с обманщиками. Иногда такие механизмы действительно вырабатываются. Часто это приводит к эволюционной "гонке вооружений": обманщики совершенствуют способы обмана, а кооператоры совершенствуют способы борьбы с обманщиками. Вот еще один пример из жизни микробов. Для бактерий Myxococcus xanthus характерно сложное коллективное поведение. Например, иногда они устраивают коллективную "охоту" на других микробов. Охотники выделяют токсины, убивающие "добычу", а затем всасывают органические вещества, высвободившиеся при распаде погибших клеток. При недостатке пищи миксококки образуют плодовые тела, в которых часть бактерий превращается в споры. В виде спор микробы могут пережить голодные времена. Плодовое тело формируется путем самосборки за счет согласованного поведения множества индивидуальных бактерий. При этом лишь часть бактерий получает прямую выгоду, а остальные жертвуют собой ради общего блага. Дело в том, что не все участники коллективного действа могут превратиться в споры и передать свои гены следующим поколениям. Многие особи выступают в роли "стройматериала", обреченного умереть, не оставив потомства. Как мы уже знаем, где альтруизм, там и паразиты-обманщики. Среди миксококков обманщики тоже есть: это генетические линии (штаммы) миксококков, не способные к образованию плодовых тел, но умеющие пристраиваться к чужим плодовым телам и образовывать там свои споры. Были проведены интересные эксперименты со смешанными культурами бактерий-альтруистов и бактерий-эгоистов. Такие культуры медленно, но верно деградируют, потому что доля паразитов неуклонно растет и в конце концов альтруистов остается слишком мало, чтобы обеспечить себя и других плодовыми телами. Но оказалось, что у миксококков в результате случайных мутаций может развиваться устойчивость к нахлебникам, то есть способность не позволять им занимать выгодные позиции в плодовом теле. Причем иногда для появления такой устойчивости достаточно одной-единственной мутации (Fiegna et al., 2006). Проблема обманщиков хорошо знакома и более сложным одноклеточным организмам, таким как социальные амебы Dictyostelium. Как и некоторые общественные бактерии, эти амебы при недостатке пищи собираются в большие многоклеточные агрегаты (псевдоплазмодии), из которых затем образуются плодовые тела. Те амебы, чьи клетки идут на построение ножки плодового тела, жертвуют собой ради товарищей, которые получают шанс превратиться в споры и продолжить род. Как и общественные бактерии, амебы страдают от социального паразитизма. У них тоже встречаются штаммы обманщиков и нахлебников. Эксперименты показали, что вероятность развития устойчивости к обманщикам в результате случайных мутаций у диктиостелиума тоже довольно высока, как и у миксококков (Khare et al., 2009). В природе идет постоянная борьба между альтруистами и обманщиками. Поэтому геномы таких организмов "настроены" естественным отбором так, что случайные мутации с большой вероятностью могут приводить к появлению защиты от той или иной разновидности обманщиков. Скорее всего, у них есть специализированные молекулярные системы "обмана" (помогающие проникать в чужие плодовые тела, не строя своих) и системы "защиты от обмана" (позволяющие опознать обманщика и не пустить его в плодовое тело). Между этими системами идет эволюционная гонка вооружений. Когда у какой-то амебы возникает полезная мутация в системе обмана, такая амеба дает начало новому штамму эффективных обманщиков. Когда у другой амебы возникнет полезная мутация в системе защиты, она даст начало штамму, защищенному от новых обманщиков. И так далее. Это очень похоже на нескончаемую гонку вооружений, идущую между патогенными микробами и генами иммунной защиты.
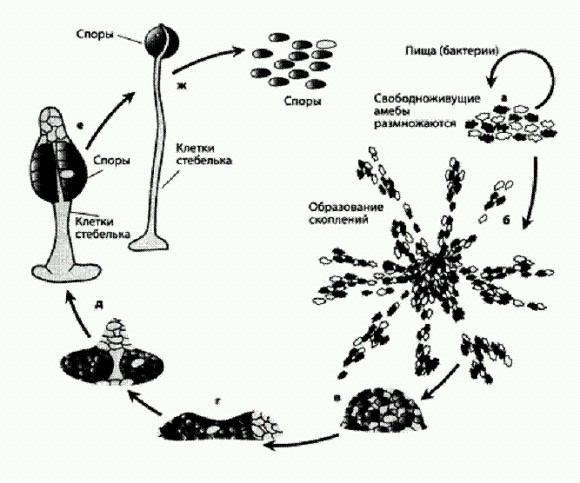
Жизненный цикл и социальный паразитизм у Dictyostelium. Темным и светлым обозначены два штамма (разновидности) амеб — "обманщики" и "честные". а — при избытке пищи амебы живут поодиночке, растут и размножаются бесполым путем (делением); половое размножение у них тоже иногда происходит, но на схеме оно не показано, б-в — при недостатке пищи амебы собираются в большие скопления, г — в результате образуются многоклеточные агрегаты длиной в несколько миллиметров, которые могут некоторое время ползать на манер слизней; их так и называют — slugs, д-ж — в конце концов многоклеточный агрегат превращается в "плодовое тело" на ножке; при этом около 20% клеток жертвуют собой, образуя ножку, а 80% превращаются в споры и получают шанс продолжить свой род. Видно, что темные клетки ("обманщики") захватили почти все лучшие места в плодовом теле и превратились в споры, предоставив всю неблагодарную работу по созданию ножки светлым клеткам ("честным"). По рисунку из Kessin, 2000. Создается впечатление, что эволюция неоднократно "пыталась" создать из социальных бактерий или простейших, умеющих собираться в плотные скопления, многоклеточный организм, но дело почему-то не пошло дальше плазмодиев и довольно просто устроенных плодовых тел. Все по-настоящему сложные многоклеточные организмы формируются иным путем — не из множества индивидуальных клеток со своими особенными геномами, а из потомков одной-единственной клетки. Это гарантирует генетическую идентичность всех клеток организма. Величина R становится равной единице, что создает идеальные условия для родственного отбора. Некоторые социальные системы, основанные на альтруизме и при этом вроде бы не защищенные от социальных паразитов, ухитряются выживать за счет разных маленьких хитростей. Недостойных, прямо скажем, высокого звания альтруиста. Например, в популяциях дрожжей одни особи ведут себя как альтруисты: они производят фермент инвертазу, расщепляющий сахарозу на легко усваиваемые моносахариды — глюкозу и фруктозу. Дрожжи могут поглощать и нерасщепленную сахарозу, но моносахариды усваиваются ими легче (то есть используются более эффективно). Некоторые дрожжевые клетки, однако, не производят инвертазу, хотя с удовольствием поедают глюкозу, добытую чужими трудами. Ведь инвертаза расщепляет сахарозу не внутри клетки, а снаружи, поэтому получившиеся моносахариды становятся доступны не только той клетке, которая произвела фермент, но и всем окружающим. Теоретически это должно было бы приводить к полному вытеснению альтруистов эгоистами. Но в реальности численность альтруистов не падает ниже определенного уровня. Дело в том, что альтруизм дрожжей при ближайшем рассмотрении оказался не совсем бескорыстным: дрожжи-альтруисты помогают всем окружающим, но 1% произведенной ими глюкозы они все-таки берут себе сразу, в обход общего котла. За счет этого однопроцентного выигрыша они, как выяснилось, могут мирно сосуществовать с эгоистами. Когда численность "эгоистов" достигает определенного (достаточно высокого) уровня, количество доступной глюкозы в популяции снижается настолько, что быть "альтруистом" становится просто-напросто выгоднее, чем эгоистом. Альтруисты начинают размножаться чуть быстрее эгоистов, и их количественное соотношение стабилизируется. Начинает работать так называемый частотно-зависимый отбор (он действует, когда приспособленность генотипа — в данном случае генотипа "альтруистов" — растет по мере снижения его частоты: ген выгоден, пока редок).
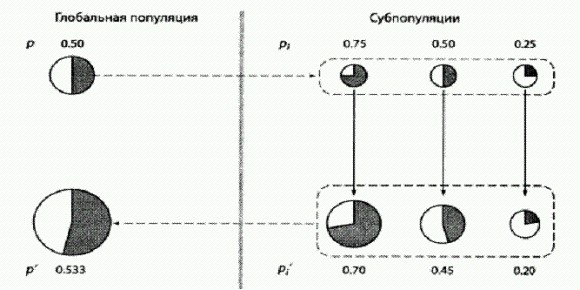
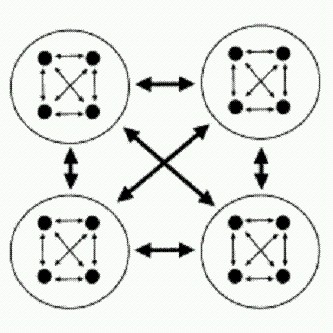
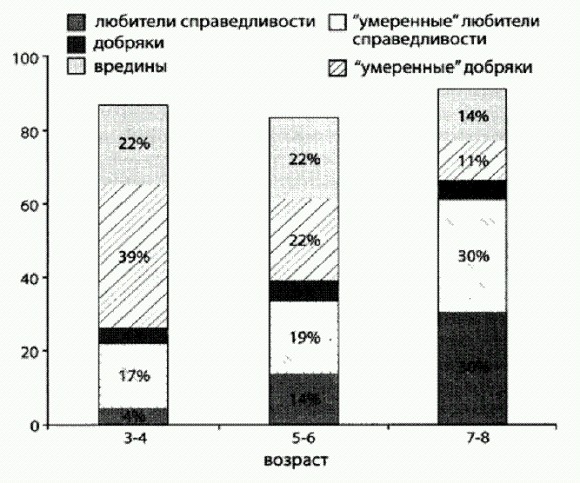
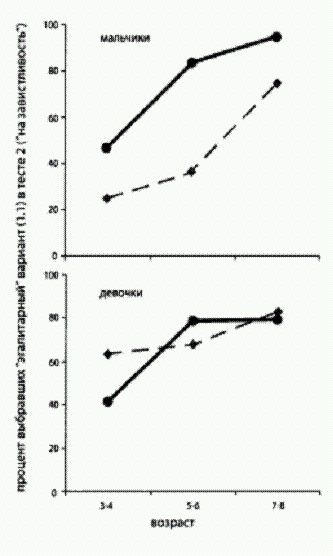
Дрожжи в последние годы стали излюбленным объектом ученых, занимающихся поведением социальных систем (на рисунке видны круглые шрамы, остающиеся на месте отпочковавшихся дочерних клеток). Но если альтруизм выгоднее эгоизма, то это уже как будто и не совсем альтруизм. Да и можно ли на таких мелких хитростях вроде жевания печенья под подушкой построить серьезную, сложную кооперативную систему? Обманщики могут быть полезны для общества? До недавних пор считалось, что положение, складывающееся в смешанной популяции дрожжей-альтруистов (производящих фермент инвертазу) и дрожжей-эгоистов (которые фермента не производят и живут на готовеньком) соответствует классической ситуации из теории игр, которая называется "игра в сугроб". Лишь в 2010 году выяснилось, что дрожжи на самом деле "в сугроб" не играют. Все оказалось сложнее и интереснее (Maclean et al., 2010). В классической "игре в сугроб" условия такие. Два игрока должны решить общую проблему (например, расчистить снежный завал на дороге или расщепить сахарозу). Если она будет решена, оба получат выигрыш b (смогут ехать дальше или получат порцию глюкозы). Чтобы проблему решить, необходимо заплатить некую цену с (например, поработать лопатой или потратить энергию на производство инвертазы). Если кооператор играет против другого кооператора, они решают проблему сообща и для каждого из них итоговый выигрыш будет равен b—с/2. Если кооператор играет против обманщика, то кооператор делает один всю работу и в итоге получает b-с, а обманщику выигрыш b достается даром. Два обманщика, играя друг против друга, ничего не делают и оба остаются с носом. Предположение о том, что дрожжи "играют в сугроб", позволило объяснить, почему в популяциях дрожжей обманщики не вытесняют кооператоров. Когда кооператоров остается слишком мало, обманщикам все чаще приходится играть друг против друга, и в итоге их стратегия становится (в среднем) менее выгодной, чем стратегия кооператоров. Однако из модели "игры в сугроб" вытекает проверяемое следствие, которое, как выяснилось, не подтверждается фактами. Состоит оно в следующем. Если дрожжи действительно "играют в сугроб", то максимальный общий выигрыш (для всей популяции "игроков" в целом) должен достигаться при полном отсутствии обманщиков в коллективе. В модели "игры в сугроб", как и в большинстве других классических моделей социальных систем, кооператоры всегда приносят коллективу только пользу, а обманщики - один сплошной вред. Иными словами, если дрожжи "играют в сугроб", то популяции дрожжей, сплошь состоящие из кооператоров, должны всегда расти быстрее, чем популяции, в которых есть обманщики. Крейг Маклин из Оксфордского университета и его коллеги решили это проверить и получили парадоксальный результат. Оказалось, что некоторая примесь обманщиков не только не вредит популяции, но и идет ей на пользу! Иными словами, в среде, где единственным источником пищи является сахароза, смешанные популяции дрожжей растут быстрее и используют ресурс эффективнее (то есть производят больше новых дрожжевых клеток на единицу съеденной сахарозы), чем популяции, состоящие из одних кооператоров. Этот результат противоречит не только модели "игры в сугроб", но и всем общепринятым представлениям о динамике социальных систем. На первый взгляд может показаться вообще невероятным, что наличие обманщиков и эгоистов, которые не производят общественно-полезного продукта, а только пользуются плодами чужих трудов, может идти на пользу коллективу. Хотя, с другой стороны, подобные ситуации были описаны и раньше (например, скорость роста колонии бактерий в присутствии антибиотика может оказаться максимальной, когда не все, а только часть бактерий вырабатывает вещество, обезвреживающее антибиотик). Чтобы разобраться в причинах парадокса, авторы разработали сложную математическую модель, призванную максимально точно отобразить все процессы и взаимодействия в исследуемых дрожжевых культурах. В модели были учтены все известные факты о биохимии, физиологии, поведении и жизненном цикле дрожжей, которые могут иметь отношение к делу. Поскольку дрожжи — классический лабораторный объект, таких фактов набралось немало. Итоговая модель представляет собой систему из 13 дифференциальных уравнений, одного взгляда на которую достаточно, чтобы повергнуть в трепет почти любого биолога, включая автора этих строк. В качестве параметров в модель были подставлены реальные цифры, полученные в ходе изучения подопытных штаммов дрожжей. Затем авторы вывели из своей модели ряд следствий, которые можно было проверить экспериментально, и все они благополучно подтвердились. В частности, модель предсказывала и тот самый парадокс, ради которого все было затеяно: модельная популяция росла на сахарозе лучше всего, если в ней помимо кооператоров были также и обманщики. Данное свойство не было заложено в модель преднамеренно, оно получилось "само" из совокупности всех известных фактов о биологии дрожжей, представленных в виде формул. Модель также предсказывала, что относительная приспособленность кооператоров (то есть эффективность их размножения по сравнению с эффективностью размножения обманщиков) должна снижаться по мере роста доли кооператоров в смешанной культуре. Иными словами, чем кооператоров больше, тем менее выгодно быть кооператором. Это предсказание было проверено экспериментально и тоже подтвердилось, причем с высокой точностью. Все это позволило авторам заключить, что модель адекватно отображает реальную ситуацию и поэтому ее можно использовать для выявления причин наблюдаемого парадокса. Анализ модели показал, что парадокс проявляется при одновременном выполнении СЛЕДУЮЩИХ ТРЕХ УСЛОВИЙ. ВО-ПЕРВЫХ, эффективность использования ресурса (в данном случае глюкозы, которая наряду с фруктозой образуется при расщеплении сахарозы ферментом инвертазой) должна снижаться по мере роста его концентрации. Иными словами, в голодные времена пища должна использоваться дрожжами более эффективно (с большим выходом биомассы на единицу съеденной глюкозы), чем в периоды изобилия. Если убрать данную зависимость из модели и сделать так, чтобы эффективность использования пищи была одинаковой при любом ее количестве, парадокс исчезает и модельные популяции, как и положено, начинают расти лучше всего при полном отсутствии обманщиков. Эксперименты подтвердили, что эффективность использования глюкозы у дрожжей действительно снижается с ростом концентрации глюкозы. Это, между прочим, означает, что величина b, то есть выигрыш, получаемый дрожжами от каждой условной единицы произведенной глюкозы, не является постоянной, как должно быть в классической "игре в сугроб", а меняется в зависимости от условий (в данном случае — от концентрации глюкозы). В результате, если в культуре очень много кооператоров, они выделяют большое количество инвертазы и производят много глюкозы сразу — так много, что эффективность использования этого ценного ресурса снижается. В условиях глюкозного изобилия дрожжи растут быстро, но неэффективно, то есть на каждый грамм съеденной глюкозы в итоге производится меньше дрожжевой биомассы, чем при более скудном рационе. Если же "разбавить" культуру кооператоров некоторым количеством обманщиков, сахароза будет переводиться в глюкозу более постепенно и в целом ресурс будет расходоваться бережнее. ВТОРОЕ НЕОБХОДИМОЕ УСЛОВИЕ состоит В ТОМ, ЧТО смешанная культура должна иметь некую пространственную структуру, то есть не быть абсолютно гомогенной. В одних областях пространства должно быть чуть больше кооператоров, в других — чуть больше обманщиков. В противном случае все ресурсы в культуре будут распределяться абсолютно поровну между всеми клетками. Модель предсказывает, что в этой ситуации тоже следует ожидать "классического" результата: максимальный групповой выигрыш будет наблюдаться при отсутствии обманщиков. Это предсказание удалось подтвердить экспериментально: если смешанные культуры очень тщательно перемешивать, парадокс исчезает, и самый быстрый рост наблюдается в культурах, на 100% состоящих из кооператоров. ТРЕТЬЕ НЕОБХОДИМОЕ УСЛОВИЕ состоит В ТОМ, ЧТО клетки не должны обладать способностью точно регулировать производство инвертазы в зависимости от наличия сахарозы в среде. В реальности дрожжи действительно производят инвертазу без оглядки на то, имеется ли в среде сахароза. Они начинают ее производить особенно интенсивно, когда им не хватает глюкозы, и делают это, даже если сахароза в среде отсутствует и от инвертазы нет никакого проку. В экспериментах дрожжи-кооператоры усиленно производили инвертазу еще долго после того, как вся сахароза была расщеплена на моносахариды. В терминах "игры в сугроб" это означает, что они продолжали разгребать снег лопатами, хотя путь уже давно был расчищен. Если в модели дать возможность дрожжам прекращать производство инвертазы, когда вся сахароза кончилась, парадокс немедленно исчезает. По-видимому, дрожжи просто не в состоянии точно определить, сколько в среде сахарозы. У них, правда, есть один рецепторный белок, реагирующий на сахарозу, но этот рецептор, к несчастью, реагирует и на глюкозу тоже. Возможно, дрожжи, как и мы, не могут определить "на вкус" концентрацию именно сахарозы, а просто чувствуют, что сладенько. Получается, что причины наблюдаемого парадокса в конечном счете сводятся к тому, что методы кооперации, практикуемые кооператорами", довольно неэффективны и негибки. Авторы предполагают, что все три условия вполне могут выполняться и в других социальных системах. Например, первое условие нам хорошо знакомо (в голодные времена пищу берегут и ею не кидаются), второе характерно для многих природных популяций (например, популяция может подразделяться на семейные группы, члены которых сходны друг с другом в среднем больше, чем с членами других групп). Третье условие с неизбежностью следует просто из того факта, что живые организмы далеко не всегда располагают всей необходимой информацией для оптимальной настройки своего поведения. Поэтому вполне возможно, что некоторая доля обманщиков может идти на пользу не только дрожжам. Однако нельзя забывать, что речь сейчас идет только о пользе для группы, а не для индивида. Причем польза для группы понимается исключительно как скорость роста этой группы (или средняя скорость размножения входящих в ее состав особей). Ясно, что для людей такое определение "пользы" далеко не всегда является адекватным. Естественному отбору, как правило, нет дела до пользы для группы Если предоставить смешанные культуры дрожжей самим себе, то под действием отбора в них установится вовсе не то соотношение кооператоров и обманщиков, при котором скорость роста группы максимальна. Ничего подобного. Установится такое соотношение, при котором "приспособленности" (скорости размножения) обманщиков и кооператоров будут равными. Или, что то же самое, относительная приспособленность тех и других будет равна единице. Это равновесное соотношение, к которому неизбежно приходит смешанная культура дрожжей под действием отбора, не совпадает с оптимальным для группы. В этом как раз и проявляется безразличие естественного отбора к нуждам коллектива. Альтруисты процветают благодаря статистическому парадоксуМогут ли быть в природе ситуации, когда альтруисты ни прямо, ни косвенно не получают никакой выгоды от своего альтруизма и совсем не умеют бороться с обманщиками, но альтруизм тем не менее развивается и процветает? Теоретически это возможно, о чем в свое время говорили и Джон Холдейн, и Уильям Гамильтон. Даже если быть эгоистом безусловно выгоднее, чем альтруистом, развитие альтруизма может идти за счет той пользы, которую получает от альтруистов вся популяция в целом, в сочетании со странным статистическим эффектом, который называется парадоксом Симпсона. В результате совместного действия этих двух факторов может возникнуть ситуация, которая интуитивно кажется невозможной: в каждой отдельной популяции процент носителей "генов альтруизма" неуклонно снижается (альтруисты всегда проигрывают в конкуренции своим эгоистичным сородичам), но если мы рассмотрим все популяции в целом, то окажется, что в глобальном масштабе процент альтруистов растет. Принцип действия парадокса Симпсона показан на рисунке. Проверить эти теоретические построения на практике довольно трудно, потому что в каждом конкретном случае, когда мы наблюдаем распространение "генов альтруизма" в природных или лабораторных популяциях, очень нелегко доказать, что здесь не замешаны ни родственный отбор, ни какие-то неизвестные нам выгоды, сопряженные с альтруизмом у данного вида живых организмов.
Гипотетический пример действия парадокса Симпсона. В исходной популяции было 50% альтруистов и 50% эгоистов (кружок слева вверху). Эта популяция подразделилась на три субпопуляции с разным соотношением альтруистов и эгоистов (три маленьких кружка справа вверху). В ходе роста каждой из трех субпопуляций альтруисты оказались в проигрыше — их процент снизился во всех трех случаях. Однако те субпопуляции, в которых изначально было больше альтруистов, выросли сильнее благодаря тому, что они имели в своем распоряжении больше "общественно-полезного продукта, производимого альтруистами (три кружка справа внизу). В результате, если сложить вместе три выросших субпопуляции, мы увидим, что "глобальный" процент альтруистов вырос (большой кружок слева внизу), р — доля альтруистов. По рисунку из Chuang et al., 2009. Чтобы выяснить, может ли парадокс Симпсона в одиночку обеспечить процветание альтруистов, американские биологи создали интересную живую модель из двух штаммов генетически модифицированных кишечных палочек (Chuang et al, 2009). В геном первого из двух штаммов ("альтруисты") был добавлен ген фермента, синтезирующего сигнальное вещество N-ацил-гомосерин-лактон (AHL), используемое некоторыми микробами для химического общения друг с другом (подобно тому, как нейроны общаются при помощи нейромедиаторов). Кроме того, в геном обоих штаммов добавили ген фермента, обеспечивающего устойчивость к антибиотику хлорамфениколу. К этому гену приделали такой промотор (регуляторную последовательность), который включает ген только в том случае, если в клетку извне поступает AHL. "Альтруисты" получили также ген зеленого светящегося белка, чтобы по силе свечения экспериментаторы могли определять процент альтруистов в популяции. "Эгоисты" ничем не отличались от альтруистов, кроме того, что у них не было гена, необходимого для синтеза сигнального вещества, и гена зеленого светящегося белка. Таким образом, сигнальное вещество, выделяемое только альтруистами, необходимо обоим штаммам для успешного роста в присутствии антибиотика. Выгода, получаемая обоими штаммами от сигнального вещества, одинакова, но альтруисты тратят ресурсы на его производство, а эгоисты живут на готовеньком. Поскольку оба штамма были искусственно созданы самими учеными и не имели никакой эволюционной истории, экспериментаторы знали наверняка, что никаких "маленьких хитростей" во взаимоотношениях альтруистов с эгоистами в их модели нет, и альтруисты не получают от своего альтруизма никаких дополнительных выгод. Кстати, авторы этой работы не пользуются "антропоморфными", по их мнению, терминами "альтруисты" и "эгоисты", а называют своих микробов производителями и не-производителями общественно полезного продукта. Ну, это дело вкуса. В среде с добавлением антибиотика чистые культуры эгоистов, как и следовало ожидать, росли хуже, чем чистые культуры альтруистов (поскольку в отсутствии сигнального вещества ген защиты от антибиотика у эгоистов оставался выключен). Однако они начинали расти лучше альтруистов, как только в среду добавляли либо живых альтруистов, либо очищенное сигнальное вещество. Альтруисты в смешанной культуре росли медленнее, потому что им приходилось тратить ресурсы на синтез AHL и бесполезного светящегося белка. Убедившись, что модельная система работает в соответствии с ожиданиями, исследователи приступили к моделированию парадокса Симпсона. Для этого они посадили в 12 пробирок со средой, содержащей антибиотик, смеси двух культур в разных пропорциях, подождали 12 часов, а затем измерили численность бактерий и процент альтруистов в каждой пробирке. Оказалось, что во всех пробирках процент альтруистов снизился. Таким образом, альтруисты во всех случаях проигрывали конкуренцию эгоистам. Однако размер тех популяций, где изначально было больше альтруистов, вырос сильнее, чем тех, где преобладали эгоисты. Когда ученые суммировали численности микробов во всех 12 пробирках, то выяснилось, что общий процент альтруистов заметно вырос: парадокс Симпсона сработал! Однако в природе никто не будет нарочно смешивать альтруистов с эгоистами в разных пропорциях и рассаживать их по пробиркам (ну или, скажем, по пещерам). Какой природный процесс может служить аналогом такой процедуры? Авторы показали, что эту роль могут играть "бутылочные горлышки" — периоды сильного сокращения численности популяции с последующим ее восстановлением. Это может происходить, например, при заселении новых субстратов очень небольшим числом микробов — "основателей". Или новых охотничьих районов небольшими группами переселенцев-гоминид. Если число основателей невелико, то среди них в силу простой случайности может оказаться повышенный процент альтруистов. Популяция, которую образует эта группа основателей, будет расти быстро, тогда как другие популяции, основанные группами с преобладанием эгоистов, будут расти медленно. В итоге парадокс Симпсона обеспечит рост "глобальной* доли альтруистов в совокупности всех популяций. Чтобы доказать действенность этого механизма, авторы смешали альтруистов с эгоистами в равной пропорции, сильно разбавили полученную культуру и стали ее высевать в пробирки порциями разного объема с приблизительно известным числом микробов в каждой порции. Размер порций оказался главным фактором, от которого зависела дальнейшая судьба альтруистов. Как и следовало ожидать, когда порции были большими, парадокс Симпсона не проявился. В большой порции, то есть в большой выборке из исходной культуры, соотношение альтруистов и эгоистов по законам статистики не может сильно отличаться от исходного, то есть 1:1. Популяции, основанные этими выборками, растут примерно с одинаковой скоростью, и альтруисты оказываются в проигрыше не только в каждой популяции по отдельности, но и во всех популяциях в целом. Однако если порции были настолько малы, что в каждой было всего несколько (не более десятка) бактерий, то среди этих порций обязательно оказывались такие, в которых альтруисты резко преобладали. Такие группы основателей давали начало быстро растущим колониям, и за счет этого общий процент альтруистов в совокупности всех популяций увеличивался. Авторы также показали, что, повторив несколько раз эту последовательность действий (разбавление культуры, расселение маленькими группами в пробирки, рост, соединение популяций в одну, опять разбавление и т.д.), можно добиться сколь угодно высокого процентного содержания альтруистов в культуре. В одном из опытов они начали со смеси, содержащей лишь 10% альтруистов, и всего за пять циклов разбавления и расселения довели их долю до 95%. Авторы указывают еще на одно условие, необходимое для распространения генов альтруизма в их модельной системе: смешанным популяциям нельзя позволять расти слишком долго. Разбавление и расселение нужно проводить до того, как растущие популяции достигнут стабильного уровня численности, заселив всю питательную среду в пробирке, потому что тогда различия по уровню численности между популяциями с разным процентным содержанием альтруистов сглаживаются, и парадокс Симпсона не может проявиться. Таким образом, естественный отбор, действуя параллельно на двух уровнях — индивидуальном и популяционном, при соблюдении определенных условий может обеспечивать развитие альтруизма даже тогда, когда в каждой отдельно взятой популяции он благоприятствует эгоистам, а альтруистов обрекает на вымирание. Полиция нравов у насекомыхПоявление многоклеточных, в том числе животных, стало крупнейшим триумфом эволюции альтруизма. В многоклеточном организме большинство клеток — это клетки-альтруисты, которые отказались от собственного размножения ради общего блага. У животных по сравнению с микробами появились новые возможности для развития кооперации, основанные на сложном поведении и обучаемости. К сожалению, те же самые возможности открылись и перед обманщиками. Эволюционная гонка вооружений продолжилась на новом уровне, и опять ни альтруисты, ни обманщики не получили решающего преимущества. Одним из важных новшеств в этой бесконечной войне стала возможность физического (а не только химического) наказания обманщиков. У некоторых социальных животных появляется институт "полиции нравов". У многих видов общественных перепончатокрылых рабочие особи физиологически вполне способны к размножению, и иногда они действительно проявляют "эгоизм", откладывая собственные яйца (неоплодотворенные, из которых могут вывестись только самцы). Однако эти яйца часто уничтожаются другими рабочими, которые таким образом "блюдут чистоту нравов" в своей колонии. Германские энтомологи решили проверить, какой из двух факторов важнее для поддержания альтруизма в обществе насекомых — закрепленное родственным отбором добровольное следование принципу разумного генетического эгоизма (гипотеза 1) или полицейский надзор (гипотеза 2). Для этого они обработали данные по 10 видам перепончатокрылых (девять видов ос и медоносная пчела). Оказалось, что чем строже "полиция нравов", тем реже рабочие совершают акты эгоизма, откладывая собственные яйца (Wenseleers, Ratnieks, 2006). Ученые проверили также влияние степени родства между рабочими в гнезде на альтруистическое поведение. Степень родства между ними часто бывает ниже идеальных 75%, поскольку царица может в течение жизни спариваться с несколькими самцами. Выяснилось, что чем ниже степень родства между сестрами-рабочими, тем сильнее полицейский надзор и тем реже рабочие ведут себя эгоистически. Это, как легко заметить, соответствует гипотезе 2 и противоречит гипотезе 1. При низкой степени родства между рабочими им становится выгоднее уничтожать яйца других рабочих. Низкая степень родства также делает более выгодным "эгоистическое" поведение, но, как видно из полученных результатов, первый фактор явно перевешивает второй. Особенности механизма наследования пола у перепончатокрылых, по-видимому, сыграли важную роль в становлении альтруистического поведения и социальности, однако у современных общественных видов альтруизм поддерживается в основном не генетической выгодой, получаемой рабочими от такого поведения, а жестким полицейским контролем. Авторы замечают, что обнаруженная ими закономерность может быть справедлива и для человеческого общества, хоть это и трудно проверить экспериментально. Общественная жизнь у людей, как и у ос, невозможна без альтруизма (индивид должен иногда жертвовать своими интересами ради общества), и в конечном счете от этого выигрывают все. Однако каждой отдельной личности во многих случаях выгодно поступать эгоистически, преследуя свои корыстные интересы в ущерб коллективу. И эффективно бороться с этим можно, к сожалению, только насильственными методами. Золотое правилоЕсли в ходе эволюции гоминид роль кооперации и альтруизма увеличивалась (а это наверняка так и было), то у наших предков непременно должны были сформироваться поведенческие и психологические адаптации, направленные на борьбу с обманщиками. Задача эволюционной психологии — их найти. Например, есть мнение (его высказал Робин Данбар, о котором говорилось в главе "Общественный мозг"), что одним из главных стимулов для развития речи у наших предков была необходимость посплетничать. Сплетни — древнейшее средство распространения компрометирующих сведений о неблагонадежных членах социума, что способствует наказанию обманщиков. Не мелковат ли повод для развития столь сложной адаптации, как речь? Нет, не мелковат. Джейн Гудолл, первая исследовательница поведения шимпанзе в природе, рассказывает, что однажды в группе шимпанзе, за которой она наблюдала, завелась самка-людоедка. То есть, простите, каннибалка. Она отбирала у других самок детенышей и пожирала их. Одна из матерей едва спасла своего ребенка и, конечно, прониклась к чудовищу сильными отрицательными эмоциями. Но что делать? Бросаться в смертный бой один на один у шимпанзе как-то не принято: ведь так можно и сдачи получить. Спустя некоторое время Гудолл наблюдала следующую сцену. Несчастная мать гуляла с несколькими дружественными самцами. Компания наткнулась на людоедку. Мать при помощи эмоциональных жестов, мимики и звуков сумела втолковать своим друзьям, что вон та дама — плохая обезьяна. Самцы поняли и устроили людоедке агрессивную демонстрацию. То есть, в переводе на русский язык, обложили в три этажа и погрозили кулаком. Все, конец истории. Никаких оргвыводов. Людоедку даже не выгнали из группы. Вряд ли такое могло случиться у вида, обладающего хотя бы примитивной речью (Бурлак, 2011). Возможность донести до соплеменников внятную сплетню может быть весьма полезной адаптацией, повышающей жизнеспособность коллектива. Одним из механизмов поддержания высокого уровня кооперации в коллективе у животных с развитым интеллектом, таких как обезьяны, может быть так называемый реципрокный, или взаимный альтруизм (Trivers, 1971). Он основан на принципе "ты мне — я тебе". Оптимальная стратегия поведения в социуме, основанном на реципрокном альтруизме, выглядит примерно так: "Помоги другому, и он в будущем поможет тебе. А если не поможет, то больше ему не помогай (еще лучше — накажи)" Реципрокный альтруизм дорог сердцу приматов вида Homo sapiens, иначе они не назвали бы "золотым правилом этики" инструкцию, содержание которой столь отчетливо перекликается с идеалом реципрокности: "Поступай с другими так же, как хочешь, чтобы поступали с тобой". Деспотизм не способствует альтруизму Швейцарские и бельгийские антропологи, наблюдавшие за поведением нескольких групп шимпанзе и бонобо в неволе, обнаружили, что уровень деспотизма в группе сильно влияет на проявления реципрокного альтруизма. Чем строже иерархия, чем выше уровень деспотизма в группе, тем реже обезьяны занимаются грумингом и делятся пищей на основе реципрокности (взаимности). В группах с эгалитарным, то есть равноправным, общественным устройством и слабо выраженной иерархией обезьяны довольно часто делятся друг с другом пищей (напомню, речь идет об обезьянах, содержащихся в неволе; в природных условиях все может быть несколько иначе). При этом наблюдаются отчетливые признаки реципрокности. Зная, что обезьяна А поделилась пищей с обезьяной Б, можно с достаточной долей уверенности предсказать, что через некоторое время обезьяна Б угостит чем-нибудь обезьяну А. Такая же реципрокность прослеживается и в груминге. Однако если группа организована более деспотично, то есть имеет жесткую многоуровневую иерархию, реципрокный альтруизм проявляется реже или вовсе отсутствует. И угощения, и груминг в этом случае имеют однонаправленный, асимметричный характер. Легко догадаться, в какую сторону направлен "поток услуг" в этой ситуации: разумеется, от подчиненных к начальству. Либо доминантные особи просто отбирают у подчиненных пищу, либо подчиненные сами с ними делятся, чтобы их умилостивить и не нарваться на начальственный гнев. Авторы делают справедливый вывод, что деспотичные варианты общественного устройства, по-видимому, не способствуют развитию реципрокного (да и любого другого) альтруизма. Ведь при деспотизме распределение благ внутри группы регулируется силовыми методами, так что для доброты и взаимовыгодного обмена места не остается. Большинство современных групп охотников-собирателей практикуют более или менее эгалитарные общественные отношения, и обычай делиться пищей с соплеменниками у них распространен чрезвычайно широко. Авторы предполагают, что в общественном устройстве наших предков — ископаемых гоминид эгалитаризм преобладал над деспотизмом. Иначе у людей вряд ли развилась бы входе эволюции столь явная наследственная предрасположенность к альтруистичному поведению (Jaeggi et al., 2010). Для реципрокного альтруизма нужны мозги. Он требует умения выделять из числа сородичей тех, кто зарекомендовал себя как эгоист, и не иметь с ними никаких дел. Тем самым достигаются сразу две цели: эгоизм оказывается "наказан" (снижается выгодность эгоистического поведения), а особь, избегающая общения с эгоистами, повышает свои шансы не быть обманутой. Исходя из этих соображений эволюционные психологи предполагают, что естественный отбор должен был выработать у наших предков специальные психологические адаптации, помогающие выявлять и запоминать обманщиков. У этой гипотезы есть проверяемые следствия. В частности, она предсказывает, что способность к запоминанию обманщиков у нас, возможно, развита сильнее, чем другие похожие способности — например, к запоминанию людей с хорошей или неизвестной репутацией. Было проведено несколько исследований с целью проверки этого предсказания. В целом оно подтвердилось; правда, при этом обнаружились неожиданные детали и появились новые вопросы. "Запоминание обманщика" складывается из двух частей: во-первых, нужно запомнить самого человека, во-вторых, что он обманщик. Это две разные задачи, которые вовсе не обязательно должны всегда выполняться одновременно и согласованно. Можно, например, запомнить лицо человека, но при этом забыть, при каких обстоятельствах мы его видели и какова его репутация. Теоретически эти два аспекта запоминания обманщиков могут быть развиты у людей в разной степени, хотя различить их в эксперименте не так-то просто. Недавно германские психологи из Института экспериментальной психологии в Дюссельдорфе показали, что люди запоминают лица обманщиков не лучше и не хуже, чем лица добропорядочных граждан. Однако информация о нечестных поступках, совершенных обманщиками, впечатывается в нашу память эффективнее, чем сведения о хороших поступках добрых людей или о нейтральных поступках лиц с неизвестной репутацией (Buchner et al, 2009). В принципе это имеет смысл, если учесть, что емкость памяти не бесконечна, а многочисленные и разнообразные социальные контакты — жизненно необходимы. Если бы мы запоминали в первую очередь плохих людей, в памяти осталось бы меньше места для запоминания тех, с кем можно иметь дело. Но если уж мы по той или иной причине запомнили какого-то человека и нам известно, что доверять ему нельзя, то очень важно поставить в памяти соответствующую "галочку", чтобы в дальнейшем по возможности с ним не связываться. В другом эксперименте те же авторы показали, что запоминание имен людей в зависимости от их репутации подчиняется той же закономерности, что и запоминание лиц. Тем самым, с одной стороны, была подтверждена выявленная ранее закономерность, с другой — получен аргумент против популярной гипотезы, согласно которой механизм запоминания обманщиков имеет особо тесную связь с системой распознавания лиц (у обезьян, включая людей, есть высокоэффективный нейронный модуль для распознавания лиц, расположенный в веретеновидной извилине височной доли) (Bell, Buchner, 2009). В этом последнем эксперименте приняли участие 193 человека (111 женщин и 82 мужчины) в возрасте от 18 до 52 лет. Тестирование проводилось индивидуально. Сначала испытуемому давали прочесть список из 36 распространенных мужских имен, причем каждое имя сопровождалось краткими сведениями о роде деятельности данного человека. Треть людей были охарактеризованы как обманщики, треть — как честные люди, об оставшейся трети сообщались нейтральные сведения, из которых нельзя было сделать вывод о моральных качествах человека. В качестве "компрометирующих" сведений использовались, например, такие истории: "Он торгует старыми автомобилями и при этом часто скрывает от покупателей информацию о серьезных дефектах своего товара". Пример положительной характеристики: "Он торгует сыром, при этом он всегда разрешает покупателям попробовать сыр и не пытается сбыть лежалый товар". Все характеристики были одинаковой длины (21 слово по-немецки), и все они ранее были испытаны в независимых тестах. Было показано, что отрицательные характеристики действительно вызывают отрицательную реакцию, положительные — положительную. Для каждого испытуемого используемые имена и характеристики комбинировались случайным образом. Участники должны были указать на основе шестибалльной шкалы, насколько им симпатичен данный человек. Как и следовало ожидать, обманщики получили низкие баллы, честные люди — высокие. На втором этапе испытуемому показывали в случайном порядке 72 имени — 36 "старых", уже знакомых ему по первому этапу тестирования, и столько же новых. Имена на этот раз не сопровождались никакими дополнительными сведениями. Испытуемый должен был указать, является ли данное имя старым или новым. Если он считал, что имя старое, то далее следовал вопрос: является ли этот человек обманщиком, честным или о его репутации нельзя сказать ничего определенного. Полученные результаты были подвергнуты статистической обработке, которая позволила расчленить акт запоминания на две составляющие: запоминание собственно имени и запоминание моральных качеств его носителя. При этом, естественно, была учтена вероятность случайного угадывания. Оказалось, что запоминание самих имен не зависит от репутации их носителей. Иными словами, имена обманщиков, честных людей и людей с неизвестной репутацией запоминались испытуемыми с одинаковой эффективностью. Однако сведения о моральном облике обманщиков запоминались лучше, чем аналогичные сведения о честных и "нейтральных" личностях. Таким образом, мы не склонны избирательно запоминать обманщиков, но если уж так получилось, что мы запомнили данного человека, то факты, порочащие его репутацию, будут запоминаться с особой тщательностью. Результаты эксперимента говорят о том, что механизм запоминания обманщиков, по-видимому, является более универсальным и менее "специфичным", чем представлялось ранее. Некоторые эксперты предполагали, что для избирательного запоминания сведений об обманщиках в мозгу существует специальный модуль, тесно связанный с системой распознавания лиц. Этому способствовало то, что до сих пор в большинстве подобных экспериментов испытуемым предлагали запоминать именно лица. Теперь, однако, стало ясно, что дело тут не в лицах — имена работают ничуть не хуже. Следовательно, если особый "модуль запоминания обманщиков" существует, он не привязан строго к системе узнавания лиц и может использовать другие персональные идентификаторы, в том числе имена. Возможно, повышенная эффективность запоминания компрометирующей информации о людях связана с тем, что такая информация вызывает у нас более сильный эмоциональный отклик (возмущение, гнев), чем сведения о хороших поступках. Такой дифференцированный эмоциональный ответ, в свою очередь, тоже может быть интерпретирован как эволюционная адаптация. Нам выгодно острее реагировать на антисоциальные поступки, чем на хорошие, и лучше запоминать их, потому что они более информативны. В человеческом обществе "хорошее" поведение (кооперативное, альтруистическое) во многих случаях просто-напросто выгоднее, чем антисоциальное (благодаря выработанным в ходе культурно-социальной эволюции законам и моральным нормам). Поэтому даже люди, от природы склонные к обману и мошенничеству, сплошь и рядом ведут себя по-честному, преследуя свои корыстные интересы, — это мало о чем говорит. Антисоциальные поступки, напротив, выдают эгоиста с головой. В других экспериментах было показано, что люди лучше справляются с разнообразными тестами и успешнее решают заковыристые задачки "на сообразительность", если условие задачи подается в контексте обмана, жульничества и нарушения моральных норм. Нам легче решить задачу, если в ней говорится, что Вася украл яблоки у Маши, а не что Маша эти яблоки сама ему подарила. Наша психика — наша созданная эволюцией душа — явно склонна проявлять повышенную чуткость к информации об обманщиках и нарушителях общественных норм. Дорогое наказание Одной из психологических адаптаций, развившихся у наших предков для противодействия социальным паразитам ("обманщикам"), по-видимому, является готовность идти на жертвы ради эффективного наказания провинившегося. Такое поведение называют "дорогостоящим наказанием" (costly punishment). Многочисленные исследования показали, что многие люди действительно готовы поступиться своими корыстными интересами ради того, чтобы как следует проучить обманщика. Это, несомненно, одна из форм альтруистического поведения, потому что человек действует в ущерб себе, но на благо обществу. "Дорогостоящее наказание" (ДН) распространено во всех исследованных человеческих популяциях. Склонность к такому поведению положительно коррелирует со склонностью к другим проявлениям альтруизма. Проблема эволюционного происхождения ДН еще недавно казалась трудноразрешимой. При помощи математического моделирования было показано, что стратегия ДН легко становится "эволюционно стабильной", то есть прочно закрепляется в популяции, если численность особей, практикующих эту стратегию, уже стала достаточно высокой. В этом случае на каждого отдельного "наказывающего" приходится в среднем мало актов ДН. Поэтому наказывающий теряет совсем немного по сравнению с теми, кто никого не наказывает, зато вся группа выигрывает сильно. Но всякий новый признак поначалу редок, то есть численность носителей новой стратегии ДН изначально должна была быть низкой. В этом случае каждому из них пришлось бы совершать очень много актов ДН, и они бы сильно проигрывали тем, кто уклоняется от участия в наказании обманщиков. Решение было найдено в 2007 году командой американских биологов и математиков. Они показали, что стратегия ДН даже при изначально низкой численности ее носителей будет быстро распространяться и обязательно станет доминирующей в модельной популяции, если сделать модель несколько более приближенной к реальности. Прежние модели допускали только три возможные стратегии: обманщики, кооператоры наказывающие и кооператоры ненаказывающие. Оказалось, что нужно добавить еще одну опцию. Модельная особь должна иметь возможность свободно выбирать, участвовать ей в рискованном коллективном мероприятии или уклониться. Например, пойти на совместную охоту на пещерного медведя или остаться дома собирать корешки и поддерживать огонь. Это будет четвертая стратегия — уклониста. На охоту пойдут представители трех остальных стратегий. При этом обе группы кооператоров будут честно рисковать здоровьем, а обманщики будут прятаться за их спинами, но после претендовать на долю добычи (уклонист от этой доли заведомо отказался — он будет кушать свои корешки). Размер добычи зависит от количества участвующих в охоте кооператоров. Риск состоит в том, что, если среди охотников окажется слишком много обманщиков, то доля добычи, доставшаяся каждому охотнику, может оказаться такой маленькой, что лучше бы он остался собирать корешки. Предполагается, что люди могут время от времени менять свой стереотип поведения, перенимая у кого-то из соплеменников его стратегию, если дела у этого соплеменника идут хорошо. Как ни странно, оказалось, что в такой модели "наказывающие кооператоры" оказываются в намного более выигрышном положении, чем при отсутствии "уклонистов". В итоге почти все особи в группе вскоре становятся "наказывающими кооператорами". Дополнительная степень свободы — добровольное, а не принудительное участие в рискованном коллективном действе, парадоксальным образом резко повышает вероятность закрепления новой разновидности альтруизма — стратегии ДН (Hauert et al., 2007). Этот еще один пример общей закономерности: эгалитарное (то есть относительно свободное и равноправное) общество создает больше предпосылок для развития альтруизма по сравнению с обществом, основанном на жесткой иерархии и деспотизме. Вопросы репутацииДалеко не все альтруистические поступки людей можно объяснить родственным отбором (мы помогаем далеко не только родственникам) или реципрокностью (мы иногда совершаем добрые поступки по отношению к людям, которые заведомо не смогут нам отплатить той же монетой). Даже не модная ныне теория группового отбора не в силах объяснить некоторые акты альтруизма, потому что никакой пользы группе они тоже не приносят. Типичный пример — добровольные пожертвования в фонд помощи голодающим детям какой-нибудь далекой страны. Казалось бы, такое поведение абсолютно бессмысленно и даже вредно как для генов жертвователя, так и для социума, к которому он принадлежит. Могут ли быть у такого поведения эволюционные корни? По-видимому, все-таки могут. Специальные психологические исследования показали, что в такой ситуации жертвователя, как ни странно, мало волнует, дойдет ли его пожертвование до адресата. Он не проявляет особого интереса к тому, насколько эффективно работает благотворительный фонд, в который он вносит деньги, и какая доля собранных средств уйдет на содержание самого фонда и накладные расходы. С другой стороны, жертвователь, как правило, хочет, чтобы о его поступке узнали окружающие — те, от чьего мнения зависит его социальный статус. А также особи противоположного пола, на которых он хотел бы произвести благоприятное впечатление. Такой механизм мотивации альтруистических поступков биологи называют непрямой реципрокностью. Выигрыш в данном случае достигается не за счет прямой отдачи по принципу "ты мне, я тебе", как при обычной реципрокности, а за счет демонстрации окружающим собственных качеств, ценимых особями противоположного пола и обществом в целом. В случае пожертвований, например, демонстрируются доброта, щедрость и материальная обеспеченность. Показные акты альтруизма — более эффективное средство саморекламы, чем "демонстративное потребление" или расточительство, о котором говорилось в главе "Происхождение человека и половой отбор" (кн. 1). Расточительство демонстрирует только материальную обеспеченность. Альтруистический акт — еще и доброту. Поэтому подобные поступки являются превосходными "индикаторами приспособленности", особенно если они достаточно щедры (дороговизна, то есть обременительность, индикатора защищает его от подделок и обеспечивает "честность сигнала", см. главу "Происхождение человека и половой отбор"). Разумеется, жертвователь должен тщательно скрывать (в том числе, желательно, и от себя самого), что он работает на публику. Нужно, чтобы окружающие подумали: "Ах, какой он добрый, щедрый и богатый". Явная показуха испортила бы весь эффект. Таким образом, склонность к демонстративным актам альтруизма, направленным на кого попало, может быть поддержана половым отбором. Особенно если дело касается вида, обладающего эффективной системой коммуникации. Сплетни, то есть, простите, эффективные механизмы распространения информации о чужих поступках избавляют альтруиста от необходимости совершать щедрые пожертвования перед носом у всех и каждого. Представляете, как бы он всем надоел и как быстро разорился! Возможно, именно благодаря наличию языка непрямая реципрокность так распространена среди людей. Но она иногда встречается и у других животных. Самый яркий пример — арабские дроздовые тимелии (Turdoides squamiceps), птицы, ведущие общественный образ жизни. Только высокоранговые самцы тимелий имеют право кормить своих сородичей. Если низкоранговый самец попробует угостить кого-то, стоящего выше него в общественной иерархии, он рискует получить взбучку. Самцы всерьез конкурируют за право совершить "добрый поступок": посидеть над гнездами в роли часового, помочь самкам ухаживать за птенцами, накормить товарища, Альтруистические акты приобрели у них отчасти символическое значение и служат для демонстрации и поддержания собственного статуса (Zahavi, 1990). Осознав важность вопросов репутации, эволюционные психологи стали организовывать эксперименты таким образом, чтобы полностью исключить любой намек на возможность какой бы то ни было реципрокности — как прямой, так и косвенной. Для этого экспериментальную ситуацию стали делать максимально "анонимной". Испытуемых всеми способами убеждали, что никто никогда не узнает об их поведении в ходе эксперимента и что ни о каких "наказании" или "награде" не может быть и речи. Однако даже в условиях полнейшей анонимности люди все равно продолжали (хотя и далеко не столь активно) совершать альтруистические поступки. Например, в тесте "Диктатор" (см. раздел "В поисках генов доброты" в главе "Генетика души") при соблюдении строгой анонимности многие испытуемые добровольно отдают часть полученных от экспериментаторов денег незнакомому и невидимому "партнеру" (которого обычно в таких экспериментах вовсе не существует, хотя испытуемый об этом не знает). В поисках причин подобных "необъяснимых" альтруистических актов психологи обнаружили, что сделать ситуацию по-настоящему анонимной в действительности не так-то просто. Похоже, люди просто не в состоянии до конца поверить, что их поступок не будет иметь никаких социальных последствий. Даже самые слабые, косвенные напоминания о возможности того, что за их действиями наблюдают, резко повышают склонность людей к альтруизму. В 2005 году американские психологи обнаружили, что люди, проходящие в условиях полной анонимности тест "Диктатор", ведут себя более просоциально (альтруистично), если на рабочем столе компьютера присутствует стилизованное изображение двух глаз (Haley, Fessler, 2005). Доходит до смешного: как показали дальнейшие исследования, достаточно разместить где-то в интерьере три точки, расположенные в виде перевернутого треугольника ('.'), чтобы испытуемые начали вести себя более альтруистично по сравнению с контрольной ситуацией, когда точки расположены наоборот (.'.). Эти результаты впоследствии были воспроизведены несколькими исследовательскими коллективами в разных странах. Полностью убрать из экспериментальной ситуации элемент непрямой реципрокности едва ли возможно. В конце концов, в реальной жизни палеолитический человек вряд ли когда-нибудь мог быть абсолютно и безоговорочно уверен в том, что о его поступке никто не узнает. И современный тоже. Какая-то доля сомнения в анонимности ситуации всегда остается, и эти сомнения, очевидно, в значительной мере подпитывают наше "врожденное нравственное чувство". Замечу к слову, что религиозные люди в экономических играх ведут себя более просоциально, если перед игрой их знакомят с текстом, где упоминается что-нибудь божественное. Точно такой же эффект на всех людей независимо от их религиозности оказывает напоминание о светских институтах, контролирующих законность и мораль (подробнее об этом мы поговорим в главе "Жертвы эволюции"). Какие мы хорошие (лирическое отступление) Подозреваю, что некоторых читателей до глубины души возмущают такие "циничные" эволюционные объяснения самых благородных и прекрасных сторон человеческого поведения. Неужели мы никогда не делаем ничего хорошего по-настоящему, искренне, без корысти и показухи? Делаем, конечно! Некоторые люди регулярно совершают добрые поступки, по-видимому, абсолютно бескорыстно и искренне, без всяких эгоистических побуждений. У таких людей даже на бессознательном уровне, скорее всего, нет эгоистических мотиваций. Человек может быть добрым "до глубины души" в полном нейробиологическом смысле этого слова. Аж до базальных ганглиев и лимбической системы. Это просто значит, что такому человеку приятно совершать добрые поступки. У нас действительно есть врожденное "нравственное чувство", на что неустанно указывали философы с незапамятных времен. Самое главное, что все это ни капельки не противоречит "циничным" биологическим теориям происхождения доброты: родственному отбору, половому отбору, реципрокному альтруизму, непрямой реципрокности. Ведь как работает эволюция поведения? Она работает путем изменения системы мотиваций, а мотивация поведения у позвоночных животных основана на эмоциях. Не на логике, тем более не на научных знаниях — на эмоциях. В этой главе речь идет о том, что у предков человека сформировалась генетически обусловленная склонность к альтруистическому поведению. Это значит, что под действием отбора закреплялись такие мутации, которые повышали вероятность того, что человеку будет приятно вести себя по-доброму в тех или иных ситуациях. Гену — выгода, нам — радость. Всю "циничную" часть работы взял на себя естественный отбор. Это он, бессовестный, ориентировался на эгоистические интересы генов. Это им руководили корысть, семейственность и реципрокность. Но он свое дело сделал. Он обеспечил нас генами, которые заставляют нейроны мозга связаться в такие сети, чтобы у нас выделялись эндорфины, когда мы делаем что-то хорошее. Поэтому нам самим вовсе не обязательно помнить об интересах генов, чтобы поступать хорошо. Мы не должны вычислять коэффициент родства, чтобы определить, за скольких племянников следует пожертвовать жизнью. Нам не обязательно помнить о реципрокности. Мы и вправду можем вовсе не думать о показухе, когда жертвуем деньги голодающим детям или сдаем кровь на донорском пункте. Нам просто приятно, и этого достаточно. Циничный естественный отбор позаботился о том, чтобы нам было приятно то, что выгодно нашим генам. Но наши чувства не становятся от этого менее настоящими. Как раз наоборот: чем циничнее он работал, тем искреннее мы сами. Мы в самом деле можем быть добрыми, щедрыми, великодушными и гуманными. По-настоящему. До самых базальных ганглиев. Склонность к альтруизму сильнее у тех, кому нечего терятьПоведенческие адаптации, связанные с альтруизмом, легче и дешевле изучать на насекомых, чем на приматах. Рассмотрим еще одно исследование, показывающее, как далек от идеала бескорыстности альтруизм общественных насекомых. У многих высокосоциальных видов рабочие особи в принципе не способны к размножению, и поэтому проблемы выбора между личными и общественными интересами для них попросту не существует. Иное дело — те виды, у которых эволюция по пути социализации и "подчинения личного общественному" не зашла столь далеко. У этих видов самка, сегодня хлопочущая над чужими детишками, завтра вполне может обзавестись своими собственными. Именно так обстоит дело у многих ос, в том числе у осы Liostenogaster flavolineata, поведение которой изучает Джереми Филд и его коллеги из Университетского колледжа в Лондоне (Field et al., 2006). Эти осы живут семьями, включающими от одной до десяти взрослых самок, из которых только одна — самая старая — откладывает яйца, а остальные заботятся о личинках. Генетический анализ, проведенный исследователями, показал, что все осы в гнезде являются родственницами, однако степень родства царицы и ее помощниц сильно варьирует. Преобладают родные сестры, но есть и кузины, и более отдаленная родня. Когда царица погибает, ее место занимает следующая по старшинству оса. Внешне помощницы ничем не отличаются от царицы, однако жизнь они ведут гораздо более тяжелую и опасную: если царица почти не покидает гнезда, то помощницам приходится все время летать за кормом для личинок, изнашивая крылышки и рискуя попасться на глаза хищнику. Неудивительно, что с переходом помощницы в ранг царицы ожидаемая продолжительность ее жизни резко увеличивается. Проблема, которую попытались решить исследователи, состоит в том, что и у этого вида, и у многих других осы-помощницы сильно различаются по степени "трудового энтузиазма". Если одни, не жалея себя, проводят в поисках пищи до 90% времени, то другие предпочитают отсиживаться в безопасном гнезде и вылетают за кормом на порядок реже. Эти различия трудно объяснить с позиций теории родственного отбора, поскольку, как показали генетические тесты, степень трудового энтузиазма помощниц не коррелирует со степенью их родства с царицей и личинками, о которых они заботятся. Ученые предположили, что каждая помощница, возможно, дозирует свой альтруизм в зависимости от того, насколько велики ее шансы стать царицей и оставить собственное потомство. Если эти шансы туманны и зыбки (как у низкоранговых молодых ос, последних в очереди на царский престол), целесообразно работать поактивнее, чтобы хоть через чужих детей передать свои гены следующим поколениям. Если же помощница имеет высокий ранг (определяемый исключительно возрастом — самая старшая из помощниц после смерти царицы занимает ее место в 90% случаев), ей выгоднее поберечь себя и поменьше рисковать. Кроме того, энтузиазм помощниц может зависеть и от размера семьи. Чем больше семья, тем больше потомства она сможет выкормить, поэтому быть царицей в большой семье выгоднее, чем в маленькой. В большой семье помощнице выгодно воздержаться от трудовых рекордов (надорвешься — много потеряешь), в маленькой — можно и поднапрячься, а если не доживешь до коронации — невелика потеря. Для начала исследователи оценили величину "трудового энтузиазма" (измеряемую как процент времени, проводимого вне гнезда) у ос разного ранга и в семьях разного размера. Выяснилось, что, в полном соответствии с ожиданиями, в маленьких семьях помощницы работают интенсивнее, чем в больших, а высокоранговые старые помощницы работают меньше, чем молодые. Затем были проведены эксперименты, позволившие вычленить влияние отдельных факторов. В первой серии экспериментов из одной семьи удаляли осу, занимающую второе место в иерархии (то есть первую по старшинству после царицы), а из другой, такой же по размерам, семьи удаляли низкоранговую молодую осу. После этого следили за поведением осы, до начала эксперимента занимавшей в иерархии третье место. В первом гнезде эта оса после удаления старшей помощницы повысила свой ранг, переместившись с третьего места на второе, во втором — осталась на третьем месте. Размер обеих семей остался одинаковым. Выяснилось, что в первом случае оса начинает работать примерно вдвое меньше, а именно столько же, сколько в контрольных (нетронутых) гнездах обычно работают осы "второго ранга". Попутно было установлено, что ключевую роль играет именно ранг, а не абсолютный возраст осы, потому что в контрольных гнездах средний возраст осы номер два составлял 116 дней, а в экспериментальных, где осы номер три были искусственно переведены на второе место, их средний возраст был равен всего лишь 55 дням. Во втором случае, когда из гнезда изымалась низкоранговая помощница, оса номер три продолжала работать столько же, сколько и раньше. Вторая серия экспериментов показала, что резкое сокращение размеров группы приводит к повышению трудовой активности помощниц даже в том случае, если не меняется ни их ранг, ни число личинок, приходящихся на одну помощницу. Исследователи в данном случае просто изымали из гнезд всех ос, кроме царицы и старшей помощницы, а также часть приплода, соответствующую доле изъятых помощниц. Оставшаяся в гнезде помощница начинала работать активнее, несмотря на то что ее шансы на "престолонаследование" нисколько не ухудшились, а число личинок, находящихся на ее попечении, вроде бы не увеличилось... (хотя тут, как мне представляется, эксперимент получился не совсем чистый. Ведь старшая помощница, как известно, самая ленивая, то есть в нетронутом гнезде она обеспечивает кормом меньше личинок, чем любая из младших помощниц. А экспериментаторы, изымая "лишних" личинок, как будто забыли об этом и оставляли ей равную долю. Если бы она продолжала работать с той же интенсивностью, что и раньше, часть личинок погибла бы с голоду) Тем не менее полученные результаты показывают, что величина "альтруистического усилия" у ос действительно регулируется в зависимости от шансов данной осы на собственный репродуктивный успех. Кому нечего терять, тот старается для общего блага, кто рассчитывает сорвать большой куш, тот себя бережет. А вот если бы мы все были клонами...Можно ли создать общество, в котором альтруизм будет поддерживаться без насилия и при этом не будет обманщиков? Ни осам, ни людям это пока не удалось. Но некоторые кооперативные системы, существующие в природе, указывают на то, что в принципе можно не допустить самого появления обманщиков. Для этого нужно свести генетическое разнообразие особей в коллективе к полному нулю. Тогда симбионты просто не смогут конкурировать друг с другом за то, кто из них ухватит себе больший кусок общего пирога. Если все симбионты генетически идентичны, эгоистическая эволюция внутри системы становится невозможной, потому что из минимального набора условий, необходимых для эволюции, — дарвиновской триады "наследственность, изменчивость, отбор" — исключается один из компонентов, а именно изменчивость. Именно поэтому эволюции так и не удалось создать нормальный многоклеточный организм из генетически разнородных клеток, но удалось создать его из клонов — потомков одной-единственной клетки. Если кооперативная система состоит из крупного многоклеточного "хозяина" и множества маленьких "симбионтов", то для хозяина самый простой путь обеспечить генетическую идентичность симбионтов — это передавать их вертикально, то есть по наследству, причем заниматься этим должен только один из полов — либо самцы, либо самки. Именно так передают из поколения в поколения свои сельскохозяйственные культуры муравьи-листорезы. При такой вертикальной передаче генетическое разнообразие симбионтов автоматически поддерживается на близком к нулю уровне за счет генетического дрейфа и "бутылочных горлышек" численности. Существуют, однако, и симбиотические системы с горизонтальной передачей симбионтов. В таких системах симбионты у каждого хозяина генетически разнородны, они сохраняют способность к эгоистической эволюции, и поэтому среди них то и дело появляются обманщики. Например, известны штаммы обманщиков среди светящихся бактерий (симбионтов рыб и кальмаров), азотфиксирующих бактерий-ризобий (симбионтов растений), микоризных грибов, зооксантелл (симбионтов кораллов). Во всех этих случаях эволюции "не удалось" обеспечить генетическую однородность симбионтов, и поэтому хозяевам приходится бороться с обманщиками иными методами, или просто терпеть их присутствие, полагаясь на те или иные механизмы, обеспечивающие баланс численности обманщиков и честных кооператоров. Все это не так эффективно, но что поделаешь: естественный отбор замечает только сиюминутную выгоду и совершенно не интересуется отдаленными перспективами. В общем, если бы не проблема обманщиков, порождаемая отсутствием у эволюции дара предвидения и заботы о "благе вида" (а не гена), наша планета, вероятно, была бы похожа на рай земной. Но эволюция слепа, и поэтому кооперация развивается только там, где то или иное стечение особых обстоятельств помогает обуздать обманщиков или предотвратить их появление. Межгрупповая вражда способствует внутригрупповому сотрудничествуЕсли у какого-то вида животных кооперация уже развилась настолько, что вид перешел к общественному образу жизни, если сформировались компактные социальные группы с относительно постоянным составом, то появляется новый фактор, который может подстегнуть развитие внутригрупповой кооперации. Этот фактор — межгрупповая конкуренция. Теперь уже не только индивиды конкурируют между собой за ценные ресурсы, но и группы. Шансы одиночек (и маленьких, слабых групп) резко снижаются. "Корыстные интересы" индивида оказываются неразрывно связаны с интересами коллектива. При острой межгрупповой конкуренции индивид может успешно размножиться, только будучи членом успешной группы. К чему это приводит, показывает модель, которую разработали американские этологи Керн Рив и Берт Холлдоблер (Reeve, Holldobler, 2007). Непосредственной целью исследователей было найти объяснение некоторым закономерностям, наблюдаемым в социальном устройстве общественных насекомых. Авторы упоминают четыре таких закономерности: 1. самая развитая кооперация (и наивысший уровень альтруизма, когда большинство особей отказывается от размножения ради заботы о чужом потомстве) характерна для коллективов, связанных близким родством; 2. у видов с большими колониями кооперация обычно развита лучше; в маленьких колониях чаще наблюдается эгоистическая "грызня" между соплеменниками за право оставить потомство; 3. у видов с мелкими и средними колониями общая производительность (отношение числа произведенных потомков к численности колонии) снижается с ростом колонии; 4. острая межгрупповая конкуренция, усиливающаяся при неравномерном пространственном распределении дефицитного ресурса, обычно коррелирует с высокоразвитой внутригрупповой кооперацией. В модели Рива и Холлдоблера каждый индивид эгоистически расходует часть "общественного пирога" на то, чтобы увеличить свою долю этого пирога. Эта потраченная на внутригрупповые склоки часть называется "эгоистическим вкладом" или "эгоистическим усилием" данного индивида. Доля, доставшаяся в итоге каждому индивиду, зависит от соотношения его собственного "эгоистического усилия" и суммы "эгоистических усилий" остальных членов группы — подобно тому как в перетягивании каната итог определяется соотношением сил, приложенных к двум концам каната. Нечто подобное наблюдается у общественных насекомых, когда они осуществляют "взаимный надзор" — мешают друг другу откладывать яйца, стараясь при этом отложить свои (см. выше). На тех же принципах строятся в модели и взаимоотношения между группами. Таким образом, получается вложенное, двухуровневое перетягивание каната. Чем больше энергии тратят индивидуумы на внутригрупповую борьбу, тем меньше ее остается для межгруппового перетягивания и тем меньше получается "общий пирог" группы. Исследуя эту модель, авторы показали, что она неплохо воспроизводит (а значит и объясняет) все четыре перечисленные закономерности. Мы не будем глубоко погружаться в математические дебри, но пару формул я все-таки приведу — хотя бы для демонстрации образа мысли авторов. Читатели, не любящие формул, могут их смело пропустить (тем более что все объясняется в тексте). Если в группе есть редкие "мутанты", отличающиеся от сородичей величиной своего эгоистического усилия, то полученная мутантом персональная доля "общественного пирога" (ресурса, добытого группой) вычисляется по формуле: S = х/(х + r (n — 1) х + (1 — r)(n-1)x*), где х — эгоистическое усилие мутанта (х = ft, где t — изначальный "запас энергии" индивида, f — доля этого запаса, которую он потратит на эгоистические цели); r — это величина из формулы Гамильтона: степень внутригруппового родства, варьирующая от 0 (нет общих генов) до 1 (все гены общие); n — число особей в группе; х* = f*t — эгоистическое усилие немутантных членов группы. Здесь учитывается, что если в группе есть мутант и есть заданная внутригрупповая степень родства (г), то соответствующая фракция всей группы (r) будет иметь "мутантный ген" и проявлять "мутантное поведение", то есть демонстрировать эгоистическое усилие, равное х, а не х*.
Вложенное перетягивание каната. Члены группы соревнуются друг с другом за свою долю "общественного пирога". Размер "пирога" в свою очередь зависит от успешности группы в соревновании с другими группами. Чем больше сил тратят особи на внутригрупповую борьбу, тем меньше их остается на общественно-полезную деятельность. По рисунку из Reeve, Holldobler, 2007. Конкурентоспособность этой группы (G) в соревновании с другими группами вычисляется как сумма всех индивидуальных "неэгоистических" усилий, направленных на рост благосостояния колонии (неэгоистическое усилие равно (1 — f) t для мутантов и (1 — f*) t для не-мутантов). Доля ресурса, отвоеванная группой в борьбе с другими группами (тот самый "общий пирог", который делят между собой члены группы), вычисляется по формуле: S = G/(G + r' (N — 1) G + (1 — r') (N - 1) G*), где r' — степень межгруппового родства, N — число групп, G* — конкурентоспособность тех групп, в которых нет мутантов. Затем, играя с этими уравнениями по законам теории игр, авторы нашли эволюционно стабильное значение f*, то есть такую величину индивидуального эгоистического усилия, при которой никакой новый мутант не сможет распространиться в популяции при данных значениях четырех переменных (N, n, r и r'). Решение получилось такое: f* = N (n — 1) (1 — r)/(Nn - 1 - г (n - 1) - (N - 1) r' (1 + r (n-1))). Величина (1 — f*) отражает интегрированность, или "сверхорганизменность" колонии. Когда 1 — f* = 0, никакой кооперации нет, и группы как таковой тоже, это полный индивидуализм. Когда величина 1 — f* близка к единице, колония становится настоящим сверхорганизмом, аналогом многоклеточного животного. Модель предсказывает, что внутригрупповая кооперация (эволюционно стабильное значение 1 — f*) должна расти с ростом внутригруппового родства (r). Это наблюдается в природе (см. выше закономерность №1) и полностью соответствует теории родственного отбора. Данный результат говорит о том, что степень родства между членами группы — отнюдь не второстепенный фактор, как считают некоторые, а мощный регулятор развития кооперации. Максимальная кооперация, согласно модели, должна наблюдаться в колониях, где большинство индивидуумов — родные братья и сестры, то есть дети одной пары насекомых-основателей (царя и царицы). "Многоцарствие" должно снижать кооперацию и способствовать внутригрупповым конфликтам, однако этот эффект, согласно модели, может быть сглажен острой межгрупповой конкуренцией, то есть большим числом конкурирующих колоний (N) (закономерность №4). Кроме того, модель предсказывает нечто весьма удивительное и из теории родственного отбора не вытекающее. Она предсказывает, что "гены кооперации" могут быть поддержаны отбором даже при полном отсутствии родства между членами группы. Из последней формулы следует, что при отсутствии родства (r = r' = 0) внутригрупповая кооперация (1 — f*) будет больше нуля при наличии хоть какой-то межгрупповой конкуренции (N > 1). Модель предсказывает, что при неизменном числе конкурирующих колоний (N) рост размера колонии (п) должен снижать кооперацию. Казалось бы, это противоречит закономерности №2. По мнению авторов, тут все дело в том, что и размер колоний, и их количество на единицу площади зависят от неравномерности распределения дефицитных ресурсов и от богатства участков, где ресурс есть. Чем богаче участок, тем обычно больше на нем колоний общественных насекомых и тем крупнее каждая колония. Таким образом, один и тот же фактор (богатство участка) положительно коррелирует с обеими переменными (n и N). Когда богатство участка растет, одновременно увеличиваются и n (что ведет к снижению кооперации), и N (что ведет к ее росту). Но в модели степень кооперации чувствительнее к изменениям N, чем n. Поэтому в итоге кооперация все-таки растет. Само по себе снижение кооперации при росте размера колонии (n) соответствует закономерности №3 и говорит о том факте, верно отраженном моделью, что в больших коллективах вольготнее чувствуют себя обманщики, паразитирующие на чужом альтруизме (в модели это "мутанты" с повышенным значением f). Авторы полагают, что их модель может быть приложима не только к насекомым, но и к другим социальным животным, включая человека. Особенно интригующим выглядит предсказание модели, согласно которому кооперация должна снижаться с ростом межгруппового родства (r'). Модель предсказывает, что кооперация должна быть развита сильнее у тех видов, у которых дочерние колонии закладываются далеко от материнских. Кроме того, межвидовая конкуренция должна лучше способствовать сплочению колонии, чем внутривидовая (так как в первом случае межгрупповое родство нулевое: r' = 0). Предельный случай альтруизма и "сверхорганизменности" у насекомых — это полная утрата рабочими особями способности к размножению. Согласно модели, это должно происходить либо при чрезвычайно острой межгрупповой конкуренции, либо при очень высокой степени внутригруппового родства. Главный вывод, который можно сделать из этой работы, состоит в том, что межгрупповая конкуренция — мощнейший стимул для развития внутригрупповой кооперации и альтруизма. Аналогии с человеческим обществом вполне очевидны. Ничто так не сплачивает коллектив, как совместное противостояние другим коллективам. Множество внешних врагов (N) — обязательное условие существования тоталитарных империй и надежное средство "сплочения" населения в альтруистический муравейник. Альтруизм, стремление к равенству и нелюбовь к чужакамВ подавляющем большинстве случаев альтруистическое поведение у животных либо направлено на близких родственников (что хорошо объясняется теорией родственного отбора), либо основано на принципе реципрокности (взаимности). По-настоящему бескорыстная забота о неродственниках в природе встречается очень редко (см. главу "В поисках душевной грани"). Люди тоже охотнее помогают "своим", чем "чужим", хотя понятие "свой" для нас не всегда совпадает с понятием "родственник". И все же, почему у людей альтруизм, как представляется, получил большее развитие, чем у других обезьян? Многие антропологи считают, что важную роль в эволюционном развитии альтруизма у наших предков могли сыграть войны. Идею о связи межгрупповых конфликтов с эволюцией альтруизма высказал еще Чарльз Дарвин в книге "Происхождение человека и половой отбор", где он написал буквально следующее: Когда два племени первобытных людей, живущие в одной стране, сталкивались между собой, то племя, которое (при прочих равных условиях) заключало в себе большее число храбрых, верных и преданных членов, всегда готовых предупреждать других об опасности и защищать друг друга, без всякого сомнения, должно было иметь больше успеха и покорить другое... Племя, обладающее перечисленными качествами в значительной степени, без всякого сомнения, распространится и одержит верх над другими племенами. Но с течением времени оно, как показывает история всех прошедших веков, будет в свою очередь покорено каким-либо другим, еще более одаренным племенем. Таким образом общественные и нравственные качества развиваются и распространяются мало-помалу по всей земле (Дарвин здесь рассуждает как групповой селекционист, то есть сторонник теории группового отбора, которая сейчас не в чести (см. выше). Многие эксперты сомневаются, что группы людей можно рассматривать как "единицы отбора". Нужно ведь еще объяснить, почему эгоисты не распространялись внутри каждой группы за счет обычного, индивидуального отбора, и почему такой индивидуальный отбор не "пересилил" гипотетический групповой, который по своей эффективности в общем случае должен сильно уступать индивидуальному. Современные модели, поддерживая основную идею Дарвина, обосновывают ее более аккуратными и изощренными аргументами, о которых пойдет речь ниже). Старая идея о связи межгрупповых конфликтов с эволюцией нравственности активно развивается в последние годы (правда, в несколько ином ключе) в рамках новой теории "сопряженной эволюции парохиального альтруизма и войн" (Choi, Bowles, 2007). Согласно этой теории, альтруизм у наших предков изначально был направлен только на членов своей группы. Такой альтруизм называют парохиальным, то есть местническим, узким, "только для своих". При помощи математических моделей было показано, что альтруизм мог развиваться только в комплексе с ксенофобией — враждебностью к чужакам. В условиях постоянных войн с соседями сочетание внутригруппового альтруизма с враждебностью к чужакам обеспечивает наибольшие шансы на выживание и успешное размножение индивидуума. Получается, что такие, казалось бы, противоположные свойства человека, как доброта и воинственность, развивались в едином комплексе: ни та, ни другая из этих черт по отдельности не способствовала бы репродуктивному успеху их обладателей. Хотя эта теория оформилась совсем недавно, выглядит она солидно и вообще очень похожа на правду. Для ее проверки психологи и антропологи уже успели провести несколько специальных исследований, с результатами которых нам будет полезно познакомиться. Швейцарские и германские психологи изучили становление альтруизма и парохиализма (предпочтения "своих") в ходе развития детей (Febr et al., 2008). В эксперименте приняли участие 229 швейцарских детей в возрасте от трех до восьми лет, среди которых не было близких родственников. Каждого ребенка просили выполнить ТРИ ПРОСТЫХ ЗАДАНИЯ. В ПЕРВОМ СЛУЧАЕ ребенок должен был выбрать один из двух вариантов: либо он сам получит конфету, а другой ребенок нет, либо обоим дадут по конфете (распределение 1,1 или 1,0). Здесь проверялось желание сделать добро другому без всякого ущерба для себя (поскольку сам испытуемый получал одну конфету независимо от сделанного им выбора). Во ВТОРОМ СЛУЧАЕ нужно было выбрать между вариантами 1,1 и 1,2. Испытуемый и на этот раз получал одну конфету независимо от принятого им решения, однако от него зависело, сколько конфет дадут другому — одну или две. Этот тест "на завистливость" был нужен, в частности, для того чтобы вскрыть истинные мотивы тех детей, которые в первом тесте выбрали вариант 1,1. Почему они "присудили" другому конфету? Те дети, которые просто желали другому добра, выберут во втором тесте вариант 1,2 (это можно интерпретировать как стремление максимизировать выгоду, получаемую другим). Те же, кто выбрал в первом тесте вариант 1,1 из-за любви к равенству и справедливости, во втором тесте тоже должны выбрать вариант 1,1. Ведь это нечестно, если мне дадут одну конфету, а ему две. В ТРЕТЬЕМ СЛУЧАЕ выбор был самым трудным: взять две конфеты себе или поделить их поровну (1,1 или 2,0). Здесь ребенок мог обеспечить ближнего конфетой только с ущербом для себя, а это уже настоящий альтруизм. Все тесты были анонимными: ребенок не знал, с кем именно ему предлагают поделиться. Ему показывали фотографию группы детей и объясняли, что конфета достанется кому-то из них. Кроме того, ребенка убеждали, что о его решении никто не узнает и поэтому не будет ни обид, ни благодарности. Таким образом ученые старались исключить любые мотивы, связанные с реципрокностью (такие как боязнь испортить с кем-то отношения), которыми может руководствоваться ребенок, делясь конфетами в реальной жизни. Чтобы выяснить ситуацию с парохиальностью, исследователи использовали два типа групповых фотографий. В одном случае на снимке были дети из того же класса или группы детского садика, что и испытуемый (ситуация "свой"). В другом случае использовали фотографию незнакомых детей (ситуация "чужой"). Такая комбинация тестов позволила получить детальную и достоверную информацию о мотивах социального поведения детей. Сначала исследователи проанализировали результаты тестов для ситуации "свой". Выяснилось, что большинство трех- и четырехлетних детей ведут себя как абсолютные эгоисты. Делая выбор, маленький ребенок обращает внимание только на то, сколько конфет достанется ему самому. Судьба анонимного "партнера" ему безразлична. В первом и втором тестах ребенок-эгоист выбирает один из двух вариантов наугад, случайным образом, а в третьем всегда берет обе конфеты себе. В соответствии с этим у трех- и четырехлетних детей частота выбора любого из двух вариантов в тестах 1 и 2 статистически не отличалась от 50%, а в тесте 3 только 8,7% детей выбрали вариант 1,1, то есть поделились с партнером. Для детей в возрасте 5-6 лет результаты первого теста оказались такими же. Во втором тесте наметилось небольшое увеличение доли детей, выбирающих вариант 1,1 (то есть тех, кто, получив одну конфету, не хочет, чтобы другому досталось две). Третий тест выявил небольшой рост числа детей, готовых поделиться с анонимным партнером (22%).
Доля пяти категорий детей в трех возрастных группах (см. пояснения в тексте). По рисунку из Fehr et al., 2008. Картина оказалась резко иной в старшей возрастной группе (7-8 лет). Почти половина (45%) детей этого возраста продемонстрировали альтруистическое поведение (поделились конфетой) в тесте 3. В тесте 1 подавляющее большинство детей (78%) выбрали вариант 1,1, то есть проявили заботу о ближнем без ущерба для себя. Тест 2 вскрыл истинные мотивы этой заботы. 80% детей во втором тесте выбрали вариант 1,1, то есть не пожелали, чтобы другому ребенку досталась лишняя конфета. Из этого авторы делают вывод, что забота о других, выявленная в тестах 1 и 3, основана не на желании сделать максимум добра ближнему, а на стремлении к равенству и справедливости. Это стремление проявляется в том, что дети отвергают "нечестную" дележку как в свою, так и в чужую пользу. До сих пор речь шла о результатах раздельного анализа трех тестов. Дополнительную информацию дало рассмотрение всех трех тестов вместе. Это позволило разделить детей на пять ГРУПП (см. рисунок): 1. "ВРЕДИНЫ", выбравшие во всех трех тестах тот вариант, при котором партнеру достается меньше всего конфет; 2. "ДОБРЯКИ", всегда выбиравшие тот вариант, при котором партнер получает максимальное число конфет; 3. дети, выбравшие наилучшие для партнера варианты в тестах 1 и 2, но отказавшиеся поделиться в тесте 3, то есть готовые делать добро лишь до тех пор, пока это не требует жертв с их стороны ("УМЕРЕННЫЕ ДОБРЯКИ"); 4. "ЛЮБИТЕЛИ СПРАВЕДЛИВОСТИ", которые всегда делят конфеты поровну; 5. "УМЕРЕННЫЕ ЛЮБИТЕЛИ СПРАВЕДЛИВОСТИ" — те, кто выбирает равную дележку в тестах 1 и 2, но поступает эгоистично в тесте 3, когда ради "торжества справедливости" пришлось бы пожертвовать конфету. Доля "вредин" среди детей от трех до шести лет составляет 22%; в возрасте 7—8 лет она сокращается до 14%. Авторы отмечают, что примерно столько же вредных личностей насчитывается и среди взрослых. Доля "добряков", как ни странно, не меняется с возрастом: таких беззаветно добрых детей оказалось около 5 % во всех возрастных группах. Доля "умеренных добряков" сокращается от 39% в возрасте 3—4 лет до 11% в 7—8 лет.
Результаты теста 2 для мальчиков и девочек в ситуациях "свой" (светлая пунктирная линия) и "чужой" (темная линия). Девочки почти не делают различий между своими и чужими, тогда как мальчики уже в 1—4 года проявляют повышенную ревность к чужакам (темная линия на верхнем графике проходит значительно выше, чем светлая). Кроме того, мальчики, особенно в раннем возрасте, заботятся о своих даже в ущерб "справедливости" (позволяют партнеру получить лишнюю конфету). Это видно из того, что светлая линия на верхнем графике в левой части проходит ниже, чем такая же линия на нижнем графике, и значительно ниже уровня 50%. По рисунку из Fehr et al., 2008. Доля "любителей справедливости" стремительно растет с возрастом: от 4% в младшей возрастной группе до 30% в старшей. Растет также и доля "умеренных любителей справедливости" (от 17% до 30%). Эти результаты заставляют задуматься. Какую роль играют в нашем обществе 5% добряков, не они ли дают нам моральные ориентиры, не на них ли держится мир? А если так, почему их всего 5%? Может быть, потому, что излишнее размножение беззаветных альтруистов создает слишком благоприятную среду для эгоистов, которые будут паразитировать на чужой доброте? Что касается "любителей справедливости", то их роль в обществе очевидна: они сдерживают развитие социального паразитизма. Интересные результаты были получены при анализе тестов, в которых "анонимным партнером" был незнакомый ребенок (ситуация "чужой"). В целом, как и следовало ожидать, дети проявляли меньше заботы о незнакомых партнерах, чем о "своих", причем это справедливо для всех возрастных групп. Самая контрастная картина получилась при раздельном анализе результатов, показанных мальчиками и девочками в тесте 2 ("проверка на завистливость"). Девочки, как выяснилось, практически не делают различий между своими и чужими в этом тесте (см. рисунок). В 3—4 года им безразлично, сколько конфет получит партнер (частота выбора любого из вариантов около 50%); в 5—6 лет начинает проявляться эгалитаризм: 70— 80% девочек выбирают вариант 1,1 и отвергают вариант 1,2, то есть не позволяют партнеру получить лишнюю конфету. Никакого предпочтения "своих" при этом не наблюдается. Мальчики, напротив, уже в 3—4 года по-разному относятся к "своим" и "чужим". "Своим" они охотно позволяют получить лишнюю конфету (около 75% мальчиков выбирают вариант 1,2). Чужакам такой подарок делают лишь около 50% мальчиков младшей возрастной группы. В 5—6 лет щедрость по отношению к своим сохраняется, тогда как по отношению к чужакам развивается сильное чувство ревности. В 7—8 лет мальчики начинают "блюсти справедливость" также и по отношению к своим, однако и в этом возрасте они присуждают чужакам лишнюю конфету намного реже, чем своим. Полученные результаты позволяют выделить стремление к равенству (эгалитаризм) как один из важнейших факторов, регулирующих социальное поведение у людей. Это свойство психики имеет наследственную природу, то есть является отчасти генетически детерминированным (Wallace et al, 2007). Нечто похожее на эгалитаризм обнаружено у американских обезьян капуцинов. Обезьян приучили выменивать лакомства на камни, которые были у них в вольере. В обмен на камешек экспериментатор давал обезьяне какое-нибудь угощение. Все шло хорошо до тех пор, пока обезьяна не замечала, что ей дали кусочек огурца, в то время как соседка за точно такой же камень получила сладкую виноградину. Обиженное животное устраивало страшный скандал и отказывалось работать (де Ваал, 2005). Полученные результаты хорошо согласуются с теорией совместного развития альтруизма и парохиализма под влиянием острой межгрупповой конкуренции. Не исключено, что эволюционная история становления этих свойств человеческой психики в общих чертах повторяется в ходе индивидуального развития детей. Характерно, что альтруизм и парохиализм развиваются у детей практически одновременно. Поскольку главными участниками межгрупповых конфликтов и войн всегда были мужчины, представляется вполне естественным, что парохиализм сильнее выражен у мальчиков, чем у девочек. В условиях первобытной жизни мужчины-воины были лично заинтересованы в том, чтобы не только они сами, но и другие мужчины группы находились в хорошей физической форме: не было смысла "блюсти справедливость" за их счет. Что же касается женщин, то в случае поражения группы в межгрупповом конфликте их шансы на успешное размножение снижались, скорее всего, не так сильно, как у мужчин (см. главу "Общественный мозг"). Разумеется, авторы исследования отдают себе отчет в том, что изученные ими свойства детской психики во многом зависят не только от генов, но и от воспитания, то есть являются продуктом не только биологической, но и культурной эволюции. Что, впрочем, не делает полученные результаты менее интересными и информативными. В конце концов, законы и движущие силы биологической и культурной эволюции отчасти сходны, а сами процессы способны плавно перетекать друг в друга. Например, новый поведенческий признак может поначалу передаваться из поколения в поколение посредством обучения и подражания, а затем постепенно закрепиться и в генах (см. главу "Генетика души"). В следующих разделах мы рассмотрим другие исследования, раскрывающие парохиальную природу человеческого альтруизма и его неразрывную связь с межгрупповыми конфликтами. Окситоцин и парохиальностьРазумеется, парохиальный альтруизм не канул в лету: он и сегодня остается весьма характерной особенностью человеческой психики и поведения. Многие люди готовы пожертвовать своими интересами (то есть совершить альтруистический поступок) ради своих. При этом они часто с не меньшей готовностью идут на жертвы и ради того, чтобы причинить ущерб представителям враждебных групп. Действия террористов-самоубийц и военные подвиги — типичные примеры такого поведения. При острой межгрупповой вражде помощь своим и вред, причиненный чужим, в равной мере идут на пользу группе. В человеческих обществах альтруистические действия обоих типов, как правило, высоко ценятся, считаются "патриотическими", "высокоморальными" и "героическими". Положительная связь между репутацией и репродуктивным успехом особей (имеются в виду те, кто выжил после геройского поступка) могла дополнительно подстегивать развитие парохиального альтруизма у наших предков (это пример непрямой реципрокности, о которой говорилось выше). Если парохиальный альтруизм так прочно укоренен в нашей психике и имеет столь глубокие эволюционные корни, то у него должна быть и вполне четкая генетическая и нейрологическая основа. Парохиальный альтруизм должен быть как-то "закодирован" в нашем геноме и в структуре мозга. Более того, вполне могут существовать специфические нейромодуляторы или гормоны, влияющие на соответствующие аспекты нашего поведения. По мере того как теория коэволюции войн и альтруизма овладевает умами специалистов, они приступают к целенаправленному поиску химических регуляторов мозговой деятельности, влияющих на проявления парохиального альтруизма. Читатели вряд ли удивятся, узнав, что первым "подозреваемым" стал окситоцин — вещество, выделяемое нейронами гипоталамуса и играющее ключевую роль в регуляции социальных и семейных отношений у животных. О влиянии окситоцина на межличностные отношения мы говорили в главе "Генетика души". Напомню, что связь окситоциновой системы регуляции социального поведения с проявлениями кооперации и альтруизма подтверждена множеством экспериментов. Установлено, что окситоцин выделяется при положительно окрашенных контактах между родственниками (например, между матерью и ребенком), влияет на доверчивость, склонность к добрым поступкам, сочувствие, помогает понимать настроение других людей по выражению лица, побуждает чаще смотреть собеседнику в глаза, повышает чувствительность к положительно окрашенным социальным стимулам (таким как добрые слова). До сих пор, однако, оставалась неизученной работа окситоциновой системы в контексте межгрупповой конкуренции. Делает ли окситоцин нас такими же добрыми и доверчивыми по отношению к чужакам, как и к своим? Это было бы весьма удивительно, ведь сотрудничество с представителями враждебной группировки ведет к ровно противоположному эффекту по сравнению с внутригрупповой взаимопомощью. Общество, как правило, расценивает такое поведение уже не как подвиг и патриотизм, а как измену и предательство. Различает ли окситоциновая система эти нюансы? По идее, она должна их различать, иначе в условиях межгрупповой вражды ее действие стало бы попросту вредным для общества и для индивидов, в него входящих. А межгрупповая вражда, скорее всего, была "нормой жизни" на протяжении почти всей человеческой истории. Некоторые установленные ранее факты подкрепляют предположение о том, что окситоцин стимулирует не альтруизм вообще, а именно парохиальный альтруизм. В частности, известно, что у некоторых животных окситоцин активизирует территориальное поведение и агрессию против "нарушителей границ". В опытах на людях было показано, что в условиях напряженного соревнования окситоцин может усиливать чувство зависти к сопернику и злорадство в случае победы. Для проверки этой гипотезы голландские психологи провели три эксперимента (De Dreu et al., 2010). Все они проводились на мужчинах (как мы помним, на женщинах влияние окситоцина не изучают, потому что окситоцин, помимо прочего, стимулирует маточные сокращения при родах). За полчаса до начала эксперимента каждый испытуемый должен был закапать себе в нос выданный ему препарат. В половине случаев это был раствор окситоцина, в половине — водичка (плацебо). В первом эксперименте всех участников разделили на команды по три человека. Команды попарно участвовали в экономической игре, основанной на классической "дилемме заключенного" (так называют ситуацию, когда каждому выгодно вести себя эгоистично вне зависимости от действий партнеров, однако совокупный выигрыш всей группы оказывается максимальным при альтруистическом поведении участников) (классическая "дилемма заключенного" выглядит примерно так. Задержаны двое подозреваемых в совершении тяжкого преступления, но у полиции недостаточно доказательств. Каждому из задержанных следователь предлагает следующие условия: "Если ты согласишься свидетельствовать против сообщника, а он будет молчать, мы тебя сразу отпустим, а его посадим на десять лет. Ему мы предложили то же самое. Если он тебя заложит, а ты будешь молчать, получишь десять лет, а его мы отпустим. Если вы оба друг друга заложите, оба получите по пять лет. Если оба будете молчать — оба отсидите по полгода и пойдете на свободу, так как у нас нет доказательств"). Каждому игроку выдали по 10 евро, и он должен был по своему усмотрению разделить эту сумму на три части. Первая часть доставалась ему целиком, вторая шла в "общественный фонд", третья — в "межгрупповой фонд". За каждый евро, внесенный в общественный фонд, все три члена команды получали по 0,5 евро. Таким образом, максимальный общий выигрыш достигается в том случае, если игроки отдадут в общественный фонд все свои деньги: тогда каждый заработает по 15 евро. За каждый евро, внесенный в "межгрупповой фонд", все члены команды тоже получали по 0,5 евро; кроме того, у каждого игрока другой команды отнимали такую же сумму. Вкладывание денег в общественный фонд рассматривалось как показатель "внутригрупповой любви". Деньги, внесенные в межгрупповой фонд, служили мерилом "межгрупповой ненависти". После введения плацебо 52% испытуемых больше всего денег оставили себе (проявили "эгоизм"), 20% самую большую сумму внесли в общественный фонд ("любовь к своим"), 28% отдали предпочтение межгрупповому фонду ("ненависть к чужим"). Под действием окситоцина только 17% участников поступили как "эгоисты", 58% проявили "любовь к своим", 25% — "ненависть к чужим". Обработав полученные результаты несколькими статистическими методами, авторы заключили, что окситоцин усиливает "внутригрупповую любовь", но не влияет на "межгрупповую ненависть". Кроме того, испытуемых попросили оценить, чего они ожидают от своих партнеров по команде и от противников в этой игровой ситуации. Их ответы позволили понять, как влияет окситоцин на доверие к своим и недоверие к чужим. Оказалось, что доверие к своим (то есть ожидание альтруистического поведения с их стороны) резко возросло под действием окситоцина. Недоверие к чужим, то есть ожидание подлости с их стороны, не изменилось. Эти результаты показывают, что окситоцин по-разному влияет на отношение к своим и чужим. Если бы люди под действием окситоцина в равной степени "добрели" по отношению ко всем окружающим, то следовало бы ожидать, что ненависть и недоверие к чужим снизятся. Но этого не произошло. Окситоцин улучшил отношение только к "своим". Это подтверждает гипотезу о том, что окситоцин стимулирует именно парохиальный альтруизм, а не альтруизм вообще. Известно, что люди сильно различаются по своей склонности к альтруизму и кооперации. Может быть, окситоцин по-разному влияет на людей с разным характером? Чтобы выяснить это, был поставлен второй эксперимент. Он отличался от первого только тем, что всех участников в начале протестировали, чтобы определить их склонность к альтруизму, и разделили по результатам тестирования на "эгоистов" и "альтруистов". Оказалось, что окситоцин влияет на обе группы одинаково: как у альтруистов, так и у эгоистов введение окситоцина усилило любовь и доверие к своим, но не повлияло на отношение к чужим. Первые два эксперимента показали, что склонность к немотивированной (не приносящей выгоды) агрессии против чужаков под действием окситоцина не усиливается. Но, может быть, окситоцин будет стимулировать агрессию, если у игроков появится возможность нажиться за счет противников? Кроме того, в первых двух экспериментах у игроков не было возможности защититься от агрессии со стороны другой команды. Может быть, окситоцин способствует принятию превентивных мер, своего рода упреждающих ударов, целью которых является защита группы от внешнего врага? Чтобы ответить на эти вопросы, был поставлен третий эксперимент,, в котором каждый игрок должен был от лица своей команды решить, будет ли он сотрудничать с командой соперников. Представитель второй команды, со своей стороны, должен был принять такое же решение. При этом игроки ничего не знали о решениях, принятых другими игроками. Распределение выигрышей строилось, как и раньше, по принципу "дилеммы заключенного", только на этот раз речь шла о сотрудничестве между командами, а не между членами одной и той же команды. Если представители обеих команд выражали готовность сотрудничать, все получали по 1 евро. Если оба представителя отказывались сотрудничать, все получали по о,6 евро. Если игрок отказывался сотрудничать, а представитель другой команды соглашался, то первая команда оказывалась в выигрыше. Величина этого выигрыша могла быть большой (члены первой команды получали по 1,4 евро) или маленькой (1,1 евро). Манипулируя этим показателем, исследователи могли выяснить, в какой степени влияет на решения игроков жадность, то есть желание обогатиться за счет противников. Если игрок соглашался сотрудничать, а представитель другой команды отказывался, то первая команда оказывалась в проигрыше, который мог быть большим (члены команды получали лишь по 0,2 евро) или маленьким (о,5 евро). Сравнение этих вариантов позволяло оценить влияние страха, или желания защитить себя и свою команду от возможных враждебных действий со стороны противников. Оказалось, что окситоцин повышает частоту "актов агрессии" по отношению к соперникам (то есть отказов от сотрудничества, что всегда ущемляло интересы противников), но не всегда, а только если такое поведение мотивировалось страхом, то есть желанием защитить группу. Окситоцин не стимулировал враждебность к чужакам под действием жадности, но стимулировал ее под действием страха. Эти выводы подтвердились результатами опросов испытуемых после игры. У участников, находившихся под действием окситоцина, выявилась повышенная готовность нанести "упреждающий удар" по соперникам, чтобы защитить команду от агрессии с их стороны. Без окситоцина стремление к таким действиям было выражено слабее. Окситоцин, однако, не усилил у игроков желание навредить соперникам из соображений "жадности", то есть для того, чтобы поживиться за их счет. Кроме того, окситоцин в этой игре, как и в первых двух экспериментах, повысил доверие к своим, но не повлиял на степень недоверия к чужим. Исследование показало, что парохиальный альтруизм действительно находится под контролем окситоциновой системы. Окситоцин улучшает отношение к своим и готовность их поддерживать и защищать, но не меняет отношения к чужакам. Окситоцин может даже стимулировать агрессию, но только в том случае, если она имеет характер "упреждающего удара" и направлена на защиту своей группы от возможных враждебных действий со стороны соперников. Окситоцин, однако, не вызывает у людей желания навредить чужакам "просто так", без всякой выгоды для себя, и не стимулирует агрессивные акты, основанные на корыстных мотивах. Иными словами, окситоцин может провоцировать "оборонительную", но не "наступательную" агрессию. Обезьяньи войны Групповое, "коалиционное" убийство себе подобных — редкое явление в животном мире. Оно характерно для ряда общественных насекомых (например, муравьев), а среди позвоночных отмечено лишь у нескольких видов: пятнистых гиен, волков, львов, африканских диких собак и некоторых приматов. Несмотря на редкость данного явления, интерес к нему очень велик, поскольку многие антропологи видят в нем истоки человеческой воинственности. Контекст смертоубийственных стычек может быть разным, но чаще всего встречаются три варианта: 1) прямое столкновение двух враждующих группировок; 2) коллективное нападение на чужака, вторгшегося на территорию группы; 3) попытка нескольких пришельцев сообща захватить власть в чужой группе. Наибольшее внимание привлекают военные рейды шимпанзе, очень похожие на примитивные военные действия многих диких (или, как сейчас принято говорить, "традиционных") человеческих племен. Небольшие сплоченные группы шимпанзе иногда вторгаются на территорию соседней группы, преследуя единственную цель: поймать и избить, порой до смерти, какого-нибудь одиноко бродящего чужака. Не щадят при этом и детенышей. Шимпанзе не страдают от избытка альтруизма, хотя бы и парохиального, поэтому "геройские" поступки им не свойственны: они предпочитают нападать только при подавляющем численном превосходстве. Дело иногда доходит до каннибализма: убитый "вражеский" детеныш может быть хладнокровно разорван на куски и съеден. Шимпанзе ценят белковую пищу, и непохоже, чтобы во время каннибальского пиршества они испытывали какие-то эмоции, кроме удовольствия от вкусной еды. По словам знаменитой исследовательницы поведения диких шимпанзе Джейн Гудолл, "животные, не относящиеся к группе, не только могут подвергнуться жестокой атаке, но и характер этой атаки может качественно отличаться от характера атаки при внутригрупповой агрессии. К жертвам нападения относятся скорее как к добыче, на которую шимпанзе охотятся; особей из других социальных групп уже не рассматривают как шимпанзе" (Гудолл, 1992). Про войны шимпанзе написано много. Гораздо меньше известно про аналогичное поведение у других обезьян. Недавно антропологи из Мексики и Великобритании, в течение многих лет наблюдающие за паукообразными обезьянами в дремучих лесах полуострова Юкатан (Мексика), обнаружили, что самцы этих приматов, подобно шимпанзе и "традиционным" Homo sapiens, периодически совершают военные набеги на чужую территорию (Aureli et al., 2006). Ученые зарегистрировали семь таких рейдов, и хотя ни одного группового убийства наблюдать не удалось, многие особенности поведения налетчиков говорят о том, что целью набегов является если не убийство, то по крайней мере устрашение иноплеменников.
Паукообразные обезьяны живут на деревьях, а воюют на земле. Наблюдения проводились в лесах на южном берегу озера Пунта Лагуна, где проживают две группы паукообразных обезьян — восточная и западная, примерно по 20—40 обезьян в каждой (численность групп многократно менялась за годы наблюдений). Вот описание одного из набегов: Утром 11 октября 2002 года мы следовали за тремя половозрелыми самцами восточной группы. В 7:45 они спустились с деревьев на землю и в полном молчании единой колонной вступили на территорию западной группы. К сожалению, через несколько минут мы потеряли их... В 8:18 мы услышали громкий лай (сигнал тревоги) в отдалении и побежали на звук. Незадолго перед тем в ту сторону направилась самка из западной группы; чужаки могли напугать ее. Прибыв на место, откуда донесся лай, мы не обнаружили самок, зато нашли самцов из восточной группы, идущих по земле (в 8:22). Мы последовали за ними. Они двигались тихо, внимательно оглядываясь и прислушиваясь, затем залезли на дерево отдохнуть и занялись грумингом, потом двинулись дальше... В 9:28 они встретили самку из западной группы, у которой, как мы знали, был грудной младенец. Самцы немедленно напали на нее. Старший и младший налетчики схватили ее и стали кусать (третий самец почти не участвовал в этом), самка громко кричала. Через несколько минут она была спасена своим взрослым сыном, внезапно появившимся на поле боя. Он прибежал по земле. Старший из разбойников напал на него, а освободившаяся мать бросилась на помощь сыну. Четыре обезьяны (два налетчика и два местных жителя) дрались около семи минут прямо перед четырьмя наблюдателями-людьми, полностью их игнорируя. В 9:35 самка и ее сын ушли прочь; самцы из восточной группы не стали их преследовать и отправились обратно на свою территорию. Младенец, которого было не видать во время драки, присоединился к своей матери лишь после ухода врагов. Как ни странно, никто не получил серьезных ранений. Самцы вернулись на свою территорию в 10:15. За весь рейд они потратили на еду лишь 3% времени. (Aureli et al., 2006). Участники рейдов всегда передвигаются по земле, хотя для паукообразных обезьян характерен древесный образ жизни, и на землю они спускаются редко. Налетчики заходят далеко вглубь вражеской территории; идут они колонной, обычно со страшим самцом во главе, хранят полное молчание и стараются не наступать на ветки и вообще производить как можно меньше шума. Они почти не проявляют интереса к съедобным плодам (которых, впрочем, вполне хватает и на их собственной территории) и редко останавливаются для отдыха. Они держатся настороже, внимательно оглядываются и прислушиваются. Обнаружив "иноплеменников", нападают, хотя в драку вступают не всегда — иногда ограничиваются тем, что прогоняют чужака. Если встретившийся чужак — самка, готовая к размножению, ее обычно не трогают или только пугают, тогда как самка, не способная в данный момент к размножению (например, кормящая мать), может подвергнуться серьезному нападению. Рейд может длиться до трех часов. Помимо чисто "террористических" целей налетчики, возможно, преследуют отчасти и амурные. В одном из рейдов старший из восточных разбойников после десятиминутной погони тихо скрылся в зарослях вместе с западной самкой; в другой раз западную самку заметили в теплой компании восточных самцов на их территории на другое утро после рейда. Все это удивительно похоже на поведение шимпанзе — с той лишь разницей, что у паукообразных обезьян в рейдах участвовали только самцы (у шимпанзе иногда к группе самцов-налетчиков может присоединиться самка), и никто из "врагов" не был убит. Впрочем, и у шимпанзе во время боевых рейдов дело редко доходит до убийства: потребовались многие годы наблюдений, прежде чем был зарегистрирован первый случай (к настоящему времени таких случаев описано 11). У менее агрессивных бонобо никто пока не наблюдал не только убийств, но даже и самих рейдов. Скорее всего, военные рейды паукообразных обезьян могут иногда заканчиваться убийством. Такое вполне могло произойти в описанном выше случае, если бы на помощь атакованной самке не подоспел ее сын. При виде налетчиков обезьяны демонстрируют сильный испуг, громко зовут на помощь — и их соплеменники действительно спешат им на выручку, с треском проламываясь сквозь заросли. Появление защитников обычно обращает разбойников в бегство. Одна из возможных причин того, что все семь рейдов закончились без жертв, состоит в том, что нападавшим ни разу не довелось наткнуться на одинокого самца. Им попадались либо самки, либо группы. Между тем шимпанзе во время боевых рейдов убивают почти исключительно самцов (10 случаев из 11). То же самое, кстати, справедливо и для людей из "традиционных" культур. "Военная активность" у шимпанзе и паукообразных обезьян зависит от числа самцов в группе и от остроты конкуренции за самок. Если самцов мало, они не предпринимают боевых рейдов и не затевают смертоубийственных драк друг с другом. Почему древесные обезьяны во время рейдов передвигаются по земле? Авторы считают, что так им легче подкрасться к врагу незамеченными. Во всех тех редких случаях, когда самцы передвигаются по земле на своей территории, они делают это с единственной целью — незаметно подкрасться к соплеменнику, на которого они собираются напасть. Ни самки, ни молодые особи никогда не совершают сколько-нибудь длительных прогулок по земле. Кстати, внутригрупповые конфликты у этих обезьян могут быть весьма жестоки: так, в апреле 2002 года, через неделю после первого зарегистрированного рейда, один из младших самцов восточной группы был убит своими сородичами (что свидетельствует о том, что паукообразные обезьяны способны к коллективному убийству). Шимпанзе тоже могут убивать "своих": например, старшие самцы могут забить до смерти молодого, если он не соблюдает субординацию и начинает держаться как высокоранговый. Что значит "держаться как высокоранговый"? Понаблюдайте за поведением обезьян в зоопарке. Даже дети Homo sapiens без труда, чуть не с первого взгляда, могут определить, кто в группе обезьян главный. Каждое движение вожака исполнено достоинства. Все как у нас, ошибиться невозможно. Раньше, когда военные рейды были известны только у человека и шимпанзе, многие исследователи полагали, что это поведение унаследовано нами от общих с шимпанзе предков (а бонобо, происходящие от тех же предков, его вторично утратили). Наличие такого же поведения у паукообразных обезьян показывает, что оно могло возникать параллельно в разных группах обезьян. Ранее также считалось, что традиция военных рейдов развилась на основе навыков коллективной охоты, имеющихся у человека и шимпанзе, тогда как для бонобо не характерно ни то, ни другое. Однако паукообразные обезьяны не знают коллективной охоты, что говорит о том, что "военные" навыки могли развиться независимо от охотничьих. Достаточно ли крови лилось в палеолите, чтобы обеспечить преимущество "генам альтруизма"?Итак, обезьяны иногда воюют, но к бурному развитию парохиального альтруизма это не приводит. Что и не удивительно: для того чтобы межгрупповая вражда стала серьезным фактором, способствующим закреплению в генофонде популяции "генов парохиального альтруизма", эта вражда должна быть достаточно острой и кровопролитной. По-видимому, на каком-то этапе своей эволюционной истории наши предки начали воевать друг с другом куда более жестоко, чем другие обезьяны. Можно предположить, что первопричиной обострения межгрупповых конфликтов стал переход от преимущественно растительной диеты к активной добыче падали в саванне. Произошло это, скорее всего, где-то на уровне хабилисов, примерно 2,3—1,8 млн лет назад. Как говорилось выше в разделе "Межгрупповая вражда способствует внутригрупповому сотрудничеству", одним из важных факторов, способствующих обострению межгрупповой конкуренции, является неравномерное распределение ресурсов. Если ресурс не "размазан тонким слоем" по обширной территории (как, например, трава в саванне или листья и в меньшей степени плоды в лесу), а встречается отдельными богатыми "пятнами" — здесь густо, а вокруг пусто, — и если к тому же этот ресурс очень ценный (питательный, как мясо) — конкуренция за такой ресурс может стать крайне острой. Мертвые туши крупных травоядных в саванне — яркий пример именно такого ресурса, очень ценного, распределенного в пространстве и времени исключительно неравномерно и большими порциями. Кровопролитные конфликты, вероятно, не прекратились и с переходом от добычи падали к охоте на крупную дичь. Иначе наследственные поведенческие признаки, закрепившиеся в связи с острой конкуренцией у падальщиков, были бы быстро, по эволюционным масштабам времени, утрачены охотниками. Можно ли как-то проверить эти домыслы? Антропологи пытаются это сделать при помощи математического моделирования, привлекая все доступные фактические данные для максимального приближения моделей к реальности. Что же показывают модели? Они показывают, что острая межгрупповая конкуренция действительно может способствовать развитию внутригруппового альтруизма и кооперации, но только при выполнении определенных условий. Условий этих несколько, но наиболее важными являются три. Во-первых, репродуктивный успех индивида должен напрямую зависеть от процветания группы (причем в понятие "репродуктивный успех" включаются любые способы передачи своих генов потомству — в том числе и через родственников, которым индивид помог выжить и которые имеют много общих с ним генов). В том, что это условие выполнялось в коллективах наших предков, сомневаться не приходится. Если группа проигрывает межгрупповой конфликт, часть ее членов погибает, а у выживших снижаются шансы вырастить здоровое и многочисленное потомство. Например, в ходе межплеменных войн у шимпанзе группы, проигрывающие в борьбе с соседями, постепенно теряют и своих членов, и территорию, то есть доступ к пищевым ресурсам. Во-вторых, как мы уже говорили, межгрупповая вражда у наших предков должна была быть достаточно острой и кровопролитной. Доказать это значительно труднее, и среди антропологов бытуют разные точки зрения на сей счет. Третье условие состоит в том, что средняя степень генетического родства между соплеменниками должна быть существенно выше, чем между группами. В противном случае естественный отбор не сможет поддержать жертвенное поведение (если исходить из предположения, что альтруизм не дает индивидууму никаких косвенных преимуществ — ни через повышение репутации, ни через благодарность облагодетельствованных соплеменников). В 2009 году Сэмюэль Боулс из Института Санта-Фе (США), один из авторов теории сопряженной эволюции парохиального альтруизма и войн, на основе имеющихся археологических и этнографических данных попытался оценить, достаточно ли сильно враждовали между собой племена наших предков и достаточно ли высока была степень родства внутри группы по сравнению с межгрупповым родством, чтобы естественный отбор мог обеспечить развитие у них внутригруппового альтруизма (Bowles, 2009). Сделав ряд правдоподобных допущений и упрощений (без которых невозможно никакое моделирование), Боулс вывел формулы, позволяющие оценить достижимый уровень развития альтруизма, то есть максимальное снижение собственного репродуктивного успеха по сравнению с соплеменниками-эгоистами, при котором естественный отбор будет благоприятствовать распространению генов, обеспечивающих такой уровень альтруизма. Боулс показал, что эта величина (с) зависит от четырех параметров: 1. К — интенсивность межгрупповых конфликтов, которую можно оценить по уровню смертности в войнах (точнее, по отношению числа погибших на войне к общему числу смертей за определенный интервал времени); 2. X — величина, показывающая, в какой степени повышение доли альтруистов (например, храбрых воинов, готовых умереть за свое племя) увеличивает вероятность победы в межгрупповом конфликте; 3. F — "коэффициент инбридинга" (инбридинг - близкородственное скрещивание), показывающий, на сколько родство внутри группы превышает родство между враждующими группами; 4. n — размер группы. Чтобы понять, в каком диапазоне могли меняться эти четыре параметра в популяциях первобытных людей, Боулс использовал множество источников данных. Интенсивность военных конфликтов у первобытных народов оценивалась на основе тщательно отобранных археологических данных по различным группам позднепалеолитических охотников-собирателей, живших от 14— 16 тыс. до 200—300 лет назад (в последнем случае это были данные по индейцам Калифорнии и Британской Колумбии). При этом использовались всевозможные поправки (например, были учтены данные, полученные в США во время войн с индейцами, согласно которым лишь около трети стрел, попавших в человека, оставляют следы на костях). Использовались также этнографические данные по тем племенам охотников-собирателей, которые еще не успели испытать на себе влияние цивилизации и не знают ни земледелия, ни животноводства. Многие из этих племен вплоть до недавнего времени вели друг с другом кровопролитные войны (особенно богатый материал такого рода есть по австралийским аборигенам). Имеющиеся данные позволяют заключить, что отношение военных потерь к общей смертности взрослого населения в палеолите колебалось в пределах от 0,05 до 0,3, в среднем — около 0,15. Иными словами, от 5 до 30% всех смертей приходилось на военные конфликты. Это чудовищно высокий уровень кровопролитности. Даже в XX веке с его двумя мировыми войнами человечеству, по-видимому, не удалось побить палеолитический рекорд. По некоторым оценкам, в XX веке умерло всего около 4 млрд 126 млн человек, из них погибло во всевозможных крупных и мелких вооруженных конфликтах не более 185 млн, то есть 4,5%. Величина F для многих популяций охотников-собирателей известна благодаря усилиям генетиков, изучающих генетическое разнообразие человечества. Приблизительные размеры человеческих коллективов в палеолите тоже известны. В итоге остается только одна величина, которую практически невозможно оценить напрямую, — X, степень зависимости военных успехов группы от наличия в ней альтруистов (героев, храбрецов). Эта величина сильно зависит от характера конфликта, тактики боя (нападение из засады или прямое столкновение), вооружения, степени численного или технического превосходства одной из сторон, и так далее. Поэтому Боулс был вынужден ограничиться очень приблизительными, "интуитивными" оценками величины X и учитывать возможность ее варьирования в очень широких пределах. Расчеты показали, что даже при самых низких значениях X естественный отбор в популяциях охотников-собирателей должен способствовать поддержанию высокого уровня внутригруппового альтруизма. "Высокий" уровень в данном случае соответствует величинам с порядка 0,02—0,03. Иными словами, "ген альтруизма" будет распространяться в популяции, если шансы выжить и оставить потомство у носителя такого гена на 2-3% ниже, чем у соплеменника-эгоиста. Может показаться, что 2-3% — это не очень высокий уровень самопожертвования. Однако на самом деле это весьма значительная величина. Боулс наглядно показал это при помощи двух несложных расчетов. Пусть изначальная частота встречаемости данного аллеля в популяции равна 0,9. Если репродуктивный успех носителей этого аллеля на 3% ниже, чем у носителей других аллелей, то уже через 150 поколений (для людей это примерно 2000—4000 лет, ничтожный срок по эволюционным меркам) частота встречаемости "вредного" аллеля снизится с 90 до 10%. Таким образом, с точки зрения естественного отбора трехпроцентное снижение приспособленности — очень дорогая цена. Теперь попробуем взглянуть на ту же самую величину (3%) с "военной" точки зрения. Альтруизм на войне проявляется в том, что воины бросаются на врагов, не щадя своей жизни, в то время как эгоисты прячутся за их спинами. Предположим, что герои-альтруисты не имеют никаких преимуществ в мирное время (что их выживаемость и число детей такие же, как у эгоистов). Все расчеты Боулса строятся на этом допущении. Предположим также для простоты, что в случае поражения в войне с соседним племенем побежденные — как альтруисты, так и эгоисты — подвергаются полному уничтожению. За победу платят своими жизнями только альтруисты. На самом деле конфликты между первобытными племенами, по-видимому, далеко не всегда заканчивались полным истреблением побежденных, но Боулс специально пересчитал имеющиеся данные по военной смертности у палеолитических охотников-собирателей в такие тотальные "войны на уничтожение". Естественно, после такого пересчета частота конфликтов оказывается низкой — реже чем раз в поколение. На основе этих допущений можно рассчитать, сколько альтруистов должно погибнуть в победоносной войне, чтобы величина с оказалась равной 0,03. Расчеты показали, что в этих условиях военная смертность среди альтруистов должна составлять свыше 20%, то есть всякий раз, когда племя сталкивается с соседями не на жизнь, а на смерть, каждый пятый альтруист должен пожертвовать жизнью ради общей победы. Надо признать, что это не такой уж низкий уровень героизма. Таким образом, уровень межгрупповой агрессии у первобытных охотников-собирателей был достаточен для того, чтобы "гены альтруизма" распространялись в человеческой популяции. Этот механизм работал бы даже в том случае, если внутри каждой группы отбор благоприятствовал исключительно эгоистам. А ведь это условие, скорее всего, соблюдалось далеко не всегда. Самоотверженность и военные подвиги могли повышать репутацию, популярность и, следовательно, репродуктивный успех людей в мирное время. Модель Боулса вполне приложима не только к наследственным (генетическим) аспектам альтруизма, но и к культурным, передающимся путем обучения и воспитания. Она даже лучше объясняет вторые, чем первые, потому что культура и традиции соседних групп охотников-собирателей обычно различаются намного сильнее, чем их гены. Нравственность основана на эмоциях, а не на рассудкеДостижения генетики, нейробиологии, экспериментальной психологии и смежных наук сегодня уже не оставляют места для сомнений в том, что человеческая нравственность имеет не только культурно-социальную, но и отчасти биологическую природу. Ясно, что отбор зафиксировал в геномах наших предков определенные мутации, склоняющие нас к просоциальному (то есть общественно-полезному) поведению. Найти эти "моральные" мутации в геноме человека генетики по-честному пытаются, но улов пока, как мы знаем, не очень велик. В ходе эволюции гоминид отбор действовал на многие гены, регулирующие развитие и работу мозга, включая систему эмоциональной регуляции поведения. Выше мы говорили о некоторых из них: ASPM, микроцефалии, FOXP2, продинорфин. Однако биологический смысл закрепившихся в этих генах мутаций выяснить трудно. Шла бы речь о мышах — давно бы все выяснили, но на высших приматах нельзя ставить генно-инженерные эксперименты (и очень хорошо, что нельзя). Пока генетики придумывают обходные маневры, можно подойти к проблеме с другого конца — нейробиологического. Для начала хорошо бы выяснить, какие отделы мозга ответственны за вынесение моральных суждений. Нейробиологи в последние несколько лет занялись этой проблемой вплотную и уже успели выяснить ряд важных фактов. Например, в 2008 году журнал Science опубликовал статью, в которой американские нейробиологи на основе данных, полученных при помощи психологических тестов, магнитно-резонансной томографии и сложной статистики, сумели разрешить один давний философский спор (пожалуй, "разрешить" — все-таки слишком громко сказано. Давние философские споры, по-моему, вообще невозможно "разрешить" до тех пор, пока они остаются философскими. Но естественные науки тем и прекрасны, что умеют перетаскивать к себе из области философии все интересные вопросы). Речь в статье идет о классической проблеме выбора между справедливостью (или равенством) и общей эффективностью (или суммарной пользой) принимаемого решения. Вот типичный пример такой дилеммы. Допустим, вы — водитель грузовика, везущий 100 кг еды жителям двух голодающих деревень. Вы можете распределить еду поровну между деревнями, но это долго, и 20 кг продуктов успеют испортиться. Однако вы можете сохранить все продукты в целости, если отдадите их жителям только одной из двух деревень. В первом случае пострадает эффективность, но будет соблюдена справедливость, а во втором — наоборот. Философы-моралисты издавна ломают копья по этому поводу. Одни считают, что превыше всего эффективность, или "суммарное количество пользы", независимо от равномерности распределения благ между людьми. Другие, наоборот, полагают, что решение, максимизирующее суммарную пользу, может быть безнравственным, если оно идет вразрез с принципами справедливости (правды, долга и т.п.) Бурные философские споры идут и по вопросу о роли эмоций в решении подобных дилемм. Одна философская школа (когнитивисты), следуя за Платоном и Кантом, подчеркивает роль рассудка. Понятие о справедливости рассматривается как результат развития абстрактного мышления и формальной логики. Другие (их называют моральными сентименталистами), следуя за Дэвидом Юмом и Адамом Смитом, полагают, что представление о справедливости уходит корнями в эмоциональную сферу и основано на сопереживании и симпатии. Первый спор трудно разрешить нейробиологическими методами — естественные науки вообще слабо приспособлены для выяснения вопроса, что хорошо, а что плохо. Их главная задача — расшифровка причин и механизмов наблюдаемых явлений, а не вынесение им моральных оценок (это относится и к эволюционной психологии. Если выясняется, что та или иная особенность нашего поведения имеет эволюционные корни, это абсолютно ничего не говорит о том, хорошим или плохим мы должны считать такое поведение сегодня. Скажем, моногамия и ксенофобия - два человеческих свойства с одинаково глубокими и прочными эволюционными корнями. При этом первое из них, по-моему, довольно хорошее, а второе - отвратительное). Зато второй спор вполне возможно разрешить естественнонаучными методами. Для этого достаточно выяснить, какие области мозга активизируются при решении этических дилемм — эмоциональные или рассудочные. Именно это и попытались сделать американские нейробиологи (Hsu et al., 2008). В эксперименте приняли участие 26 добровольцев (17 женщин и девять мужчин) в возрасте от 29 до 55 лет, здоровые и имеющие высшее образование. Испытуемым было предложено принять ряд решений по распределению денежных средств, выделенных на питание детей в одном из детских домов Уганды. Участников ознакомили с именами и краткими биографиями 60 детей-сирот, причем и детский дом, и дети были настоящие. Испытуемые имели возможность сделать денежное пожертвование в пользу детей, и они пожертвовали в среднем по $87. Затем испытуемым было объявлено, что решения, которые они примут в ходе тестирования, будут реально учтены при распределении денег между детьми. Для наглядности все денежные суммы переводились в количество порций еды по нынешним угандийским ценам (сумма, необходимая для пропитания одного ребенка в течение месяца, составляет сегодня в Уганде пять долларов США). Тесты были составлены таким образом, чтобы испытуемые, выбирая один из двух вариантов распределения средств, вынуждены были каждый раз делать выбор между эффективностью и справедливостью. В первым случае голодным детям доставалось в сумме больше еды, но распределялась она между детьми очень неравномерно. Во втором — дележка была более честная, но суммарное количество еды уменьшалось. Одни испытуемые чаще выбирали эффективность, другие — справедливость. Пока они проходили тесты, работа их мозга регистрировалась при помощи ФМРТ. Главный смысл работы состоял в том, чтобы выявить участки мозга, избирательно реагирующие на отдельные составляющие этической дилеммы, обозначенные исследователями как эффективность, справедливость и общая польза (интегральная оценка качества достигнутого результата, основанная на совместном учете двух предыдущих показателей). Чтобы вычленить эти компоненты и оценить их количественно, результаты тестирования были подвергнуты сложной статистической обработке. Для каждого испытуемого был вычислен коэффициент а , отражающий степень неприятия несправедливости (или стремления к равенству). Затем для каждого теста были вычислены следующие показатели: 1) эффективность принятого решения, точнее, достигнутый выигрыш в эффективности (dM) — суммарное количество порций, доставшихся детям в выбранном варианте, по сравнению с количеством порций, которое досталось бы им в другом варианте; 2) "проигрыш в справедливости" (dG) — степень неравномерности распределения порций между детьми в выбранном варианте по сравнению с отвергнутым вариантом; 3) общая польза (dU = dM — adG, где a — средняя степень неприятия несправедливости испытуемыми). Затем на основе данных ФМРТ были выявлены участки мозга, активность которых коррелирует со значениями этих показателей. Вот тут-то и обнаружилось самое интересное и неожиданное. Для каждого из трех показателей нашелся один (и только один!) маленький участок мозга, активность которого в ходе тестирования коррелировала со значением данного показателя. Для общей пользы (dU) таким участком оказалось хвостатое ядро. Эффективность принятого решения (dM) коррелировала с активностью скорлупы. Для dG — меры несправедливости — ключевым участком мозга оказалась кора островка, область, связанная с чувством эмпатии (сопереживания). Похоже, здесь же формируется и чувство справедливости. Корреляция между dG и активностью коры островка наблюдалась на этапах обдумывания ситуации и принятия решения. Наблюдая за активностью коры островка, можно было предсказать, какое решение примет испытуемый: высокая активность предвещала решительный выбор в пользу справедливости (и в ущерб эффективности). Все три выявленные области мозга ассоциируются с эмоциональной, а не рассудочной сферой психики. Это, конечно, не значит, что этические решения принимаются без участия рассудка. Рассудок, надо полагать, тоже в этом участвует, но активность соответствующих рассудочных областей мозга оказывается примерно одинаковой в разных ситуациях и не зависит от того, какой будет сделан выбор—в пользу эффективности или справедливости. Рассудок, может быть, что-то там просчитывает, но окончательное решение, по-видимому, строится все-таки на основе эмоций. Авторы рассматривают полученные результаты как весомый довод в пользу моральных сентименталистов. Ясно, что решение этических дилемм неразрывно связано с эмоциями, причем эмоциональная оценка эффективности, справедливости и общей пользы принимаемого решения осуществляется, как выяснилось, тремя разными отделами мозга. При принятии решения происходит нечто вроде взвешивания баланса сил (то есть эмоций) или судебного разбирательства: кора островка защищает интересы справедливости, а скорлупа "голосует" за эффективность. Баланс противоречивых желаний подводит хвостатое ядро. По той же схеме, путем сопоставления сигналов от разных участков — "представителей" спорящих сторон, мозг, по-видимому, принимает решения и в других спорных ситуациях, например когда нужно сопоставить риск и возможный выигрыш. Что касается хвостатого ядра и прилегающей к нему области прозрачной перегородки (septum pellucidum), то новые результаты хорошо согласуются с данными, полученными ранее, согласно которым этот участок мозга играет важную роль в интеграции различных психических "переменных", в вынесении "социально ориентированных" оценок, а также в таких явлениях и поступках, как доверие к другим людям или жертвование на благотворительные нужды. Мораль и отвращениеЧто ж, приходится признать, что многие наши моральные оценки и суждения основаны больше на эмоциях, чем на рассудке. Но хорошо ли это? Могут ли эмоции служить надежной основой благополучного существования и развития социума? Ведь это не обязательно должно быть так. В ходе эволюции гоминид древняя система эмоциональной регуляции поведения подверглась определенным модификациям, направленность которых мы можем определить исходя из чисто теоретических соображений. Эти модификации должны были повышать репродуктивный успех людей каменного века. Они были выгодны генам хабилисов, эректусов, гейдельбержцев и ранних сапиенсов. Вовсе не факт, что они выгодны генам современных горожан. Тем более самим горожанам. Тем более обществу, которое эти горожане образуют. Эволюция идет медленно, она не может поспеть за быстро меняющейся жизнью. Поэтому большинство адаптаций у всех живых существ ориентированы не на нынешние условия, а на условия более или менее далекого прошлого (и хорошо, если с тех пор условия не сильно изменились). К тому же адаптации всегда ориентированы на интересы генов, а не организмов. Если у нас есть врожденная склонность считать какие-то поступки хорошими, то единственный "высший смысл", стоящий за этой склонностью, вполне может состоять в том, что генам эректусов было почему-то выгодно, чтобы эректусам нравились такие поступки. Гены эректусов от этого лучше тиражировались. На здоровье, но при чем тут мы? Однако до сих пор не только дилетанты, но и некоторые эксперты твердо убеждены, что эмоции, интуитивные побуждения и прочие природные позывы — вполне надежные критерии истинности в этических вопросах. Эта точка зрения основана на предположении, или, скорее, на интуитивной убежденности в том, что первая, непосредственная, не затуманенная всяческими умствованиями эмоциональная реакция и есть самая верная, потому что она идет "из глубины души" и несет в себе "глубинную мудрость". Голос сердца, одним словом. На это особенно напирают противники клонирования, стволовых клеток, искусственного осеменения и других технологий, "покушающихся на самое святое" и "вызывающих естественное отторжение". Тем временем дотошные нейробиологи проникают все глубже в пресловутые "глубины души", и то, что они там находят, не всегда похоже на мудрость, которую следует почитать превыше рассудка. Несколько исследовательских коллективов в последние годы активно изучают природу отвращения — одной из базовых человеческих эмоций, которая, как выясняется, сильно влияет на общественную мораль и социальные отношения (Jones, 2007). Нельзя сказать, чтобы отвращение было исключительно человеческим чувством: другим животным оно тоже свойственно, но в меньшей степени и в гораздо более простых формах. И обезьяна, и кошка, и новорожденный младенец, взяв в рот что-нибудь неприятное на вкус, могут выплюнуть это с характерной гримасой. Но от "невкусно" до "противно" немалая дистанция. Только человек, вышедший из младенческого возраста, способен отказаться от пищи лишь на том основании, что она не там лежала или не к тому прикасалась. Пол Розин из Пенсильванского университета, один из пионеров в данной области исследований, полагает, что по мере развития разума первичная, унаследованная от далеких предков эмоция резко расширилась, включив в себя, в частности, идею контакта, перенесения "скверны" путем прикосновения. Так, добровольцы, участвовавшие в экспериментах Розина, наотрез отказывались пить сок, к которому прикоснулся усиком простерилизованный таракан, или есть из безупречно чистого ночного горшка. Из этой особенности первобытного мышления, очевидно, выросла так называемая контагиозная магия (См.: Фрезер Дж. "Золотая ветвь", глава 3. http://www.psylib.ukrweb.net/books/freze01/txt03.htm). У других животных и новорожденных детей ничего подобного не наблюдается. Биологический, эволюционный смысл отвращения более-менее понятен: это вполне адаптивное, способствующее выживанию стремление избежать контактов с заразой, не есть негодную и опасную пищу, а также сохранить собственную целостность, удерживая внутри то, что должно быть внутри (например, кровь), и снаружи то, что должно быть снаружи (например, фекалии). Отвращение у людей отчетливо делится на "первичное" — это практически бессознательная психическая реакция на всякие мерзости — и "вторичное", или моральное, касающееся более абстрактных предметов, таких как идея клонирования. Связь между ними самая тесная. Во всех без исключения человеческих культурах принято распространять слова и понятия, обозначающие объекты первичного отвращения, на людей, нарушающих моральные и общественные нормы, — например, на лживых политиков и продажных чиновников. Люди, заклейменные таким образом, могут даже восприниматься как источник некоей мистической "заразы" вроде каких-нибудь тараканов. К примеру, предложение надеть на себя хорошо выстиранный свитер Гитлера не вызывает у большинства людей ни малейшего энтузиазма. По мнению Розина, это означает, что идея "заразности" в человеческом сознании распространяется и на моральные качества личности, иначе чем объяснить неприязнь к ни в чем не повинному свитеру. Психолог Пол Блум из Иельского университета настроен более скептически: по его мнению, настоящее отвращение люди испытывают только к тем абстрактным идеям, которые непосредственно ассоциируются с объектами первичного отвращения, а во всех остальных случаях (например, когда говорят об "отвратительных политических технологиях") это не более чем метафора. Джонатан Хайдт из Вирджинского университета полагает, что нашел доказательство единой физиологической природы первичного и морального отвращения: ему удалось экспериментально показать, что обе эмоции приводят к замедлению пульса, а при особенно острой реакции — еще и к чувству "комка в горле". По мнению Хайдта, это показывает, что моральное отвращение — никакая не метафора, а самое настоящее отвращение. Бразильский нейробиолог Жоржи Молл пришел к сходным выводам, наблюдая за активностью мозга испытуемых при помощи ФМРТ. Оказалось, что при первичном и моральном отвращении возбуждаются одни и те же области мозга, а именно боковая и средняя орбитофронтальная кора, — эти области отвечают и за некоторые другие неприятные переживания, такие как сожаление об упущенных возможностях. Однако выявились и различия: моральное отвращение связано с более сильным возбуждением передней части орбитофронтальной коры, которая считается более эволюционно молодой и, по-видимому, отвечает за обработку наиболее абстрактных эмоциональных ассоциаций. Независимо от того, являются ли первичное и моральное отвращение одним и тем же или разными чувствами, первичное отвращение само по себе может оказывать вполне реальное влияние на наши моральные суждения и оценки и, как следствие, на наше отношение к людям и социальное поведение. Психологи из Принстонского университета при помощи ФМРТ показали, что возбуждение отделов мозга, отвечающих за страх и отвращение, снижает активность тех отделов, которые отвечают за жалость, сочувствие и вообще за восприятие других людей как людей (в отличие от неодушевленных объектов). Иными словами, вид противного грязного бомжа автоматически вызывает чувство отвращения, которое мешает нам помыслить об этом человеке как о личности, заставляя воспринимать его как "кучу мусора". Отвращение, шерсть и блохи Шимпанзе и другие наши ближайшие родственники не ночуют две ночи подряд на одном месте, поэтому с гигиеной у них нет проблем и для выживания им не обязательно испытывать чувство отвращения ко всяким пакостям. Они могут с легким сердцем нагадить поутру в собственное гнездо, где провели ночь, потому что возвращаться сюда никто не собирается. Когда у ранних Homo в связи с изменением способов добычи пропитания и разделением труда между полами появились постоянные базы, где люди подолгу жили в тесном контакте друг с другом, эти проблемы встали во весь рост. Поначалу несчастные предки, скорее всего, тонули в отбросах и дурели от расплодившихся вшей. Результатом, по-видимому, стали три крупных эволюционных изменения: 1) появился новый вид насекомых — человеческая блоха, Pulexirritans. Вши есть и у нечеловеческих обезьян, а вот блохи бывают только у животных с постоянным логовом, потому что большая часть жизни блохи проходит не на теле жертвы, а среди всякой трухи на полу жилища; 2) люди приобрели чувство отвращения, наводившее их обезьяньи мозги на правильные с гигиенической точки зрения мысли; 3) они утратили большую часть волосяного покрова, чтобы ограничить поле деятельности вшей (хотя, конечно, есть и другие гипотезы происхождения человеческой безволосости). Розин, Хайдт и некоторые их коллеги предполагают, что отвращение может играть существенную — и в основном негативную — роль в жизни человеческих коллективов. Если изначально отвращение выполняло в основном функции гигиенического характера, то в ходе дальнейшей эволюции это чувство, похоже, было "рекрутировано" для выполнения совсем иных, чисто социальных задач. Объект, вызывающий отвращение, должен быть отброшен, изолирован или уничтожен, от него необходимо дистанцироваться. Это делает отвращение идеальным сырым материалом для развития такого мощного механизма механизма поддержания целостности группы, как ксенофобия. Сплоченность группы повышала ее шансы на выживание, а противостояние внешним врагам — наилучший способ добиться максимальной сплоченности. Возможно, еще на заре человеческой истории наши предки научились испытывать отвращение к разного рода чужакам, "не нашим", "не таким, как мы". Известный психолог из Гарвардского университета Марк Хаузер (Его книга "Мораль и разум" издана на русском языке в 2008 году) отмечает, что непростые отношения между группами бывают не только у человека, но и у других общественных животных, которые тоже прекрасно умеют отличать своих от чужих. Но люди почему-то особенно сильно зациклены на своих межгрупповых различиях и придают им по сравнению с другими животными непомерно большое значение. Для подчеркивания межгрупповых различий сплошь и рядом привлекаются морально-нравственные оценки, в том числе основанные на чувстве отвращения (к примеру, русское слово "поганый" исходно значило всего-навсего "иноверец, язычник"). По словам Хайдта, если первичное отвращение помогало выжить индивидууму, то моральное отвращение помогало выжить коллективу, сохранить целостность социума — "и именно здесь отвращение проявляет себя с самой отвратительной стороны". Политики во все времена активно использовали отвращение как инструмент сплочения и подчинения коллективов, натравливания одних групп на другие. Нацистская пропаганда называла евреев крысами и тараканами. Те же эпитеты применяли к своим противникам инициаторы массовой резни в Руанде в 1994 году. Если люди начинают испытывать отвращение к чужакам, они уже не могут воспринимать их как людей, чувствовать жалость или сострадание. По мнению Молла и других экспертов, отвращение и по сей день служит источником предвзятости и агрессии. Нужно десять раз подумать, прежде чем принимать решения на основе подобных эмоций, идущих "из глубины души". История это подтверждает. Были времена, когда омерзительными, нечистыми считались, к примеру, женщины (особенно во время менструации), психически неполноценные люди или межрасовый секс. Сегодня мало кто в цивилизованных странах будет защищать подобные взгляды, и многие действительно — на физическом уровне — перестали испытывать отвращение ко всему перечисленному. Если в прошлом отвращение не было хорошим моральным индикатором, то с чего бы ему стать таковым сегодня? Во многих случаях то, что кажется нам отвратительным, действительно плохо и вредно, но это не значит, что цивилизованные люди должны строить свои отношения на психологических пережитках далекого эволюционного прошлого. Известно, что люди сильно различаются по степени выраженности эмоции первичного отвращения: одни чуть в обморок не падают при виде таракана или неспущенной воды в унитазе, а другим хоть бы что. Блум и его коллеги обнаружили значимую корреляцию между этим показателем и политическими убеждениями. Люди, склонные испытывать сильное отвращение к "первичным" стимулам, чаще придерживаются консервативных взглядов и являются убежденными противниками клонирования, генно-модифицированных продуктов, гомосексуализма, мини-юбок, искусственного осеменения и прочих безобразий. Люди с пониженной брезгливостью, напротив, обычно имеют либеральные взгляды и просто не могут понять, отчего все вышеперечисленное может кому-то казаться отвратительным. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||