|
||||
|
|
ГЛАВА 6. ВЕЛИКОЕ РАССЕЛЕНИЕ САПИЕНСОВПервые шагиВ предыдущих главах мы упоминали, что, судя по генетическим данным, современное внеафриканское человечество в основном происходит от небольшой популяции выходцев из Африки. В основном — потому что, как показали недавние открытия в области палеогенетики, наши африканские предки в процессе своего расселения по Евразии все-таки скрещивались как минимум с двумя популяциями евразийских аборигенов — неандертальцами и загадочными денисовцами. Гибридизация с неандертальцами произошла вскоре после выхода из Африки и еще до того, как сапиенсы начали заселять обширные просторы Евразии. Поэтому неандертальские гены есть у всех современных евразийцев примерно в одинаковом количестве (около 2,5 %). Денисовские гены есть только у меланезийцев. Это значит, что скрещивались с денисовцами лишь отдельные группы сапиенсов, потомки которых впоследствии осели в Новой Гвинее и на островах к востоку от нее. Некоторые генетические данные указывают также на возможность скрещивания восточноазиатских сапиенсов с тамошними реликтовыми эректусами (Сох et al., 2008), но эти выводы, скорее всего, будут оставаться "предварительными" до тех пор, пока не удастся прочесть геном какого-нибудь позднего азиатского эректуса. До сих пор, однако, остаются неясными время и маршрут выхода сапиенсов из африканской прародины. Генетические данные не позволяют датировать это событие со сколько-нибудь приемлемой точностью. Археологические данные тоже, к сожалению, довольно сомнительны. Древнейшими свидетельствами присутствия "анатомически современных людей" за пределами Африки считаются находки в пещерах Схул и Кафзех на севере Израиля. Там обнаружены скелеты людей современного типа, хотя и с некоторыми архаичными чертами, сближающими их с древнейшими сапиенсами из Эфиопии (см. главу "От эректусов к сапиенсам"). Возраст костей сапиенсов из пещер Схул и Кафзех — 119 ± 18 и 81 ± 13 тыс. лет. Это примерно совпадает с последним межледниковьем, когда климат на Ближнем Востоке был относительно теплым и влажным. До начала этой теплой эпохи в данном районе, возможно (есть некоторые сомнения в датировках), проживали привычные к холоду неандертальцы. Скорее всего, чтобы попасть в Палестину, сапиенсы пересекли засушливые районы Северной Африки по "Нильскому коридору". Но этот первый исход из Африки, похоже, не имел далеко идущих последствий, потому что в период между 65 и 47 тыс. лет назад в том же районе регистрируется присутствие уже не сапиенсов, а снова неандертальцев. Возможно, неандертальцы пришли сюда с севера в связи с похолоданием и оттеснили теплолюбивых сапиенсов обратно в Африку. Многие эксперты полагают, что современное внеафриканское человечество в основном происходит не от той популяции, которая проникла в Азию "северным путем" по Нильскому коридору около 120 тыс. лет назад, а от другой группы выходцев из Африки, которая покинула родной материк "южным путем", то есть перебралась через Баб-эль-Мандебский пролив (глубокий и опасный для мореплавателей пролив с сильными течениями, соединяющий Красное море с Аденским заливом Индийского океана. Название пролива в переводе означает "ворота слёз" (или скорби)) в Южную Аравию и затем расселялась вдоль побережья Индийского океана на восток, оставляя на своем пути характерные раковинные кучи. Хронология этого второго исхода из Африки остается дискуссионной и опирается в основном на косвенные данные. Некоторые эксперты полагают, что он состоялся около 85 тыс. лет назад, и уже вскоре после этого сапиенсы добрались до Индонезии и южного Китая. Эта версия представлена в интерактивной презентации The Journey of Mankind (http://www.bradshawfoundation.com/journey/), основанной на трудах Стивена Оппенгеймера из Оксфордского университета. Одна из книг Оппенгеймера, "Изгнание из Эдема. Хроники демографического взрыва", переведена на русский язык (2004). Однако бесспорные следы присутствия сапиенсов (то есть ископаемые кости анатомически современных людей) в южной и Юго-Восточной Азии начинаются лишь примерно с 42 тыс. лет назад (хотя уже 48—43 тыс. лет назад сапиенсы проникли в Австралию, см. ниже). Ряд археологических находок, подтверждая косвенные данные генетики, указывает на более раннее (начиная примерно с 80 тыс. лет назад) присутствие людей на южном побережье Азии. Но как доказать, что это именно сапиенсы, а не какие-нибудь денисовцы или иные потомки более древних мигрантов из Африки? Ведь находят, как правило, только каменные орудия. Например, на юге Индостана непосредственно под толстым слоем вулканического пепла, оставленного грандиозным извержением вулкана Тоба 74 тыс. лет назад, обнаружены каменные орудия, сходные с африканским средним палеолитом (Petraglia et al., 2007). Самое удивительное, что точно такие же орудия присутствуют и непосредственно над слоем пепла! Получается, что древние индийцы сумели пережить катастрофу: отсиделись где-то, а потом как ни в чем не бывало продолжили заниматься своими делами на старом месте. С точки зрения некоторых экспертов, такая жизнестойкость является важным аргументом в пользу того, что это были именно сапиенсы. В начале 2011 года журнал Science опубликовал новые археологические данные, подтверждающие очень ранний выход сапиенсов из Африки "южным путем". К сожалению, человеческих костей по-прежнему нет: речь снова идет лишь о каменных орудиях, но все-таки эта находка значительно повышает достоверность реконструкций Оппенгеймера и его единомышленников и даже позволяет привязать пересечение Баб-эль-Мандеба к еще более ранней дате. Международная команда археологов из Великобритании, Объединенных Арабских Эмиратов, США, Украины и Германии сообщила о результатах раскопок палеолитической стоянки, расположенной на юго-востоке Аравийского полуострова, неподалеку от Ормузского пролива (соединяющего Персидский залив с океаном) (Armitage et al., 2011). Древние люди жили в небольшом гроте у подножия каменистой возвышенности Джебель-Файя. Каменная кровля грота к настоящему времени обвалилась. Раскопки в Джебель-Файя проводились в 2003—2010 годах. Сверху залегают культурные слои железного и бронзового веков, под ними — неолитический слой, затем идет чистый песок без признаков присутствия человека, а еще ниже находятся три среднепалеолитических слоя: А, В и С. На самом деле есть еще четвертый слой D, самый древний, но о нем авторы ничего конкретного пока не сообщили: не успели как следует изучить. Из того, что изучено, интереснее всего слой С. Его возраст, по результатам оптико-люминесцентного датирования, составляет от 127 ± 16 до 95 ± 13 тыс. лет. Самое главное, что каменные орудия из этого слоя как по своей "типологии" (форме, размерам, соотношению орудий разных типов и другим формальным признакам), так и по технологиям обработки камня очень похожи на изделия, производившиеся в это время сапиенсами восточной Африки. В Леванте, где селились то сапиенсы, то неандертальцы, каменная индустрия в те времена сильно отличалась от того, что найдено в слое С в Джебель-Файя. По мнению археологов, создатели каменной индустрии слоя С почти наверняка были выходцами из Восточной Африки. То есть, конечно, сапиенсами, потому что в Африке в то время других людей не было.
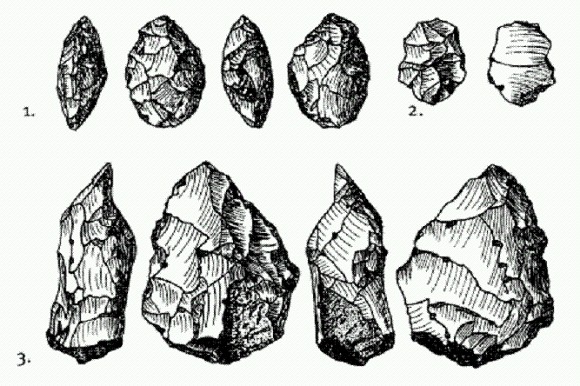
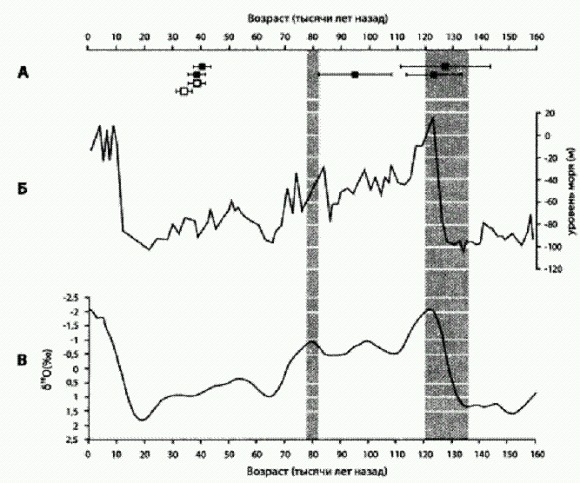
Орудия из слоя С в Джебель-Файя (127—95 тыс. лет назад). 1 — листовидное обоюдоострое орудие, 2 — леваллуазский отщеп, 3 — заготовка для обоюдоострого ручного рубила (бифаса). По рисунку из Armitage et al., 2011. Слой С без перерыва переходит в В, о возрасте которого в статье не сообщается (наверное, не смогли определить). Затем после перерыва, представляющего собой прослойку песка без орудий, идет слой А возрастом 38—40 тыс. лет. Еще выше — опять чистый песок, продолжающийся вплоть до появления неолитических орудий возрастом 10—9 тыс. лет. Каменные изделия слоев В и А — среднепалеолитические, похожие на орудия из слоя С, но отличающиеся от того, что находят на других ближневосточных и африканских стоянках тех времен. Эти данные в переводе с археологического языка означают примерно следующее. По-видимому, сапиенсы впервые появились в Джебель-Файя в начале последнего (рисс-вюрмского) межледниковья, около 130—125 тыс. лет назад. Это был на редкость удобный момент для преодоления Баб-эль-Мандебского пролива и миграции вдоль южного побережья Аравии. Вплоть до примерно 130 тыс. лет назад, пока длилось предпоследнее (рисское) оледенение, уровень моря был очень низким — на 100 м ниже, чем сейчас. Пересечь сузившийся и обмелевший пролив можно было и раньше, однако до тех пор, пока не началось глобальное потепление, климат в южной Аравии был слишком засушлив, чтобы люди могли здесь долго продержаться. Потом, когда климат улучшился, уровень моря поднялся, и пролив расширился. Однако в самом начале межледниковья, по-видимому, был краткий период, когда море еще не успело подняться, а климат в Южной Аравии уже стал благоприятным для людей. Это видно на рисунке: кривая уровня моря (Б) около 130 тыс. лет назад еще не начала подниматься, а влажный период, обозначенный серой вертикальной полосой, уже начался.
А — оптико-люминесцентный возраст трех проб из слоя С (черные квадратики справа) и двух проб из слоя А (черные квадратики слева), а также двух проб из Лстерильногоя слоя, залегающего над слоем А (белые квадратики). Б - уровень Красного моря. В — колебания глобальной температуры (о них судят по содержанию тяжелого изотопа в морских отложениях): чем выше кривая, тем теплее климат. Вертикальными серыми полосами показаны периоды наиболее благоприятного, влажного климата в Южной Аравии. По горизонтальной оси — время в тысячах лет назад. По Armitage et al., 2011. Не исключено, что часть южноаравийской популяции сапиенсов двинулась дальше на восток и дала начало всему внеафриканскому человечеству. Может быть, они успели пересечь и Ормузский пролив до подъема уровня моря (собственно, в то время пролива там не было, Персидский залив был сушей, нужно было пересечь только большую реку, образованную слиянием Тигра и Евфрата). О том, каким образом в их генофонд попали неандертальские гены, пока можно лишь гадать: возможно, были какие-то древние миграции между южной Аравией и Левантом. Так или иначе, на восток ушли не все. Об этом свидетельствуют палеолитические слои В и А. Люди, оставившие каменные орудия в этих слоях, по-видимому, жили в Джебель-Файя уже в относительной изоляции, потому что никакие технологические новшества, в том числе верхнепалеолитические технологии, к ним не проникали ни из Африки, ни из Леванта. Они сохраняли разнообразные приемы обработки камня, характерные для слоя С и африканского среднего палеолита, но кое-что, видимо, позабыли: в слоях В и А отсутствуют орудия, изготовленные при помощи леваллуазской технологии, хорошо знакомой людям слоя С (а также другим среднепалеолитическим сапиенсам и неандертальцам). Тем временем по мере развития последнего, Вюрмского, оледенения климат в Аравии становился все более засушливым. 40 тыс. лет назад процесс зашел слишком далеко, территория опустынилась, и люди исчезли из этого района вплоть до начала следующего (нынешнего) межледниковья. Лишь около 10 тыс. лет назад здесь снова появилось население, но это уже были обладатели совсем другой, несравненно более высокой неолитической культуры. Впрочем, если все эти увлекательные теории верны, неолитические пришельцы (как и все мы, внеафриканские сапиенсы) должны быть потомками древнейших обитателей южной Аравии. Открытие АвстралииИ все же следует помнить, что данные о начальных этапах расселения сапиенсов вдоль южных берегов Азии по-прежнему скудны и во многих случаях сомнительны. По-настоящему массовое и бесспорное присутствие сапиенсов в различных точках Евразии регистрируется начиная с 50 тыс. лет назад. Примерно в это время сапиенсы сумели перебраться из юго-восточных районов Азии в Австралию. Это подтверждается как археологическими, так и генетическими данными — в частности, анализом мтДНК австралийских аборигенов. Так, в 2006 году антропологи и генетики из Австралии, Великобритании, Швеции и Швейцарии сообщили в журнале American Journal of Physical Anthropology о результатах анализа мтДНК 69 аборигенов из разных районов Австралии (van Holst Pellekaan et al., 2006). Имеющиеся у современных людей варианты мтДНК подразделяются на три "макрогруппы": Li, L2, L3, причем первые две встречаются только в Африке. Макрогруппа L3 в свою очередь делится на ряд групп. Две из них (М и N) были у тех людей, которые вышли из Африки и потомки которых заселили все остальные материки. Поэтому все неафриканское человечество относится к двум большим митохондриальным линиям — М и N, и австралийцы не являются исключением. Все известные на сегодняшний день варианты мтДНК австралийцев распадаются на пять групп: AuB, AuA, AuC, AuD, AuE, причем первая группа принадлежит к линии М, а четыре другие — к N. Все пять групп, по-видимому, встречаются только в Австралии. Анализ подтвердил, что Австралия была заселена очень давно (не менее 40 тыс. лет назад). Это соответствует археологическим данным, согласно которым люди жили в Австралии уже 43—48 тыс. лет назад. В то время австралийцы вовсе не были отсталым народом. Возле озера Манго (Новый Южный Уэльс) обнаружены два уникальных древних захоронения, мужское и женское. Тело мужчины было покрыто красной охрой, а женщина, по-видимому, была кремирована. Это древнейшие в истории человечества документированные случаи использования кремации и охры в погребальных обрядах. Возраст обоих захоронений — около 40 тыс. лет, но люди в этом районе появились, вероятно, на 6-10 тыс. лет раньше (Bowler et al., 2003). Культура первых австралийцев, по-видимому, оставалась на среднепалеолитическом уровне. Характерные для верхнего палеолита инновации появлялись в Австралии постепенно, намного позже, чем в Европе, Африке и Азии, и далеко не в полном объеме (Habgood, Franklin, 2008). Однако 50— 45 тыс. лет назад верхнего палеолита как такового не существовало еще нигде. Вскоре после появления людей в Австралии произошло массовое вымирание почти всех крупных местных животных. Многие исследователи связывают это событие с негуманными методами охоты древних австралийцев. Впрочем, не исключено, что вымирание мегафауны в действительности протекало не столь стремительно и растянулось на десяток тысячелетий. Число первопоселенцев, вероятно, было довольно велико, и они были достаточно разнообразны в генетическом отношении. Этим объясняется, в частности, присутствие в Австралии обеих "глобальных" линий — М и N. Некоторые признаки указывают на родство первопоселенцев Австралии с древними обитателями Новой Гвинеи. Данные по мтДНК не дают оснований предполагать, что в более поздние эпохи имели место новые волны переселения в Австралию из других регионов (или в обратном направлении). Однако анализ Y-хромосом австралийцев показал, что в течение последних 10 тыс. лет в Австралию проникло какое-то количество выходцев из Индии. Несоответствие результатов по мтДНК и Y-хромосоме может объясняться множеством причин. Например, пришельцы из Индии могли быть мужчинами, а если с ними и были женщины, они могли не оставить прямых потомков по женской линии. Похоже, австралийцы оставались генетически изолированными от остального человечества большую часть своей истории. Более того, значительная степень изоляции имелась также и внутри самой Австралии между разными ее регионами. Это подтверждается резко неравномерным распределением генетических вариаций по территории Австралии. Кажется удивительным, что после первой, очень древней и, по-видимому, многочисленной волны переселенцев Австралия оказалась отрезанной от мира на десятки тыс. лет, и никто (или почти никто) больше не переселялся туда даже из соседних регионов, таких как Новая Гвинея. Все эти выводы носят предварительный характер. Для их уточнения необходимо собрать гораздо больше материала, а это не так-то просто. Некоторые социально-политические и культурологические факторы в настоящее время препятствуют массовому сбору необходимых анализов (проб ДНК) у коренных австралийцев. Возможно, аборигены до сих пор не простили тех европейских ученых, которые 100—150 лет назад на полном серьезе предполагали, что коренные жители Австралии и Тасмании могут оказаться искомыми "переходными формами между обезьяной и человеком". Изменения климата не были причиной массового вымирания австралийских животных В период Великого расселения сапиенсов по планете прокатилась волна массового вымирания крупных животных. В Старом Свете и Америке множество видов крупных зверей исчезло 10—15 тыс. лет назад, в Австралии это произошло раньше — около 40 тыс. лет назад. Характерно, что и в обеих Америках, и в Австралии вымирание произошло вскоре после прихода туда людей. Многие специалисты полагают, что главной причиной вымирания была деятельность первобытных охотников; другие отводят ведущую роль переменам климата. Например, гибель экосистемы мамонтовых степей (10—12 тыс. лет назад) связывают с потеплением и таянием вечной мерзлоты, а катастрофу австралийской биоты, произошедшую на 30 тыс. лет раньше и сопровождавшуюся опустыниванием большей части материка, — с аридизацией климата (предполагают, что ранее на Австралию проливалось больше дождей). В 2007 году австралийские палеонтологи получили новые данные, заставляющие усомниться в правомочности "климатической" гипотезы (Prideaux et al., 2007). Исследователи описали уникальный комплекс ископаемых плейстоценовых животных из трех пещер на юге Австралии. Эти пещеры долгое время служили своеобразными естественными ловушками: животные падали в них сквозь небольшие отверстия и либо сразу разбивались о каменный пол, либо не могли выбраться из ловушки и погибали от голода. Возраст ископаемых костей и вмещающих отложений, определенный при помощи множества независимых методов, составляет от 100 до 400 тыс. лет. Пещеры получили условное название Thylacoleo caves ("пещеры сумчатого льва"). В пещерах обнаружены остатки 69 видов позвоночных, из которых 33 вымерли, не дожив до конца плейстоценовой эпохи. Количественное соотношение представителей разных групп животных в палеокомплексе (например, малое число древесных листоядных форм и высокое разнообразие ящериц) свидетельствует о том, что плейстоценовый климат в этом районе Австралии был, как и сегодня, довольно засушливым, однако растительность была намного разнообразнее. Такие же результаты дал и анализ изотопного состава зубной эмали ископаемых животных (о климате можно судить по содержанию изотопов 13С и 18О). Сегодня в этом районе простираются сухие кустарниковые степи с низким разнообразием растительной и животной жизни. В плейстоцене здесь тоже было сухо, но вместо однообразных кустов росли пышные леса. Хотя растительных остатков в пещерах не найдено (из-за особых условий захоронения там не сохранилась даже пыльца), о богатстве растительности свидетельствуют, например, кости нескольких видов древесных кенгуру, а также очень высокое разнообразие всевозможных растительноядных сумчатых, масса тела которых варьировала ОТ 4 ДО 200 кг. Таким образом, климатические изменения не могли стать причиной резкого обеднения флоры и фауны Южной Австралии в конце плейстоцена, поскольку климат, судя по всему, существенно не менялся. Авторы полагают, что причиной экологической катастрофы могли стать пожары. Деревья, которые предположительно могли здесь расти, отличаются повышенной пожароопасностью из-за большого количества выделяемых ими эфирных масел. К аналогичным выводам относительно причин опустынивания центральной Австралии пришел и другой коллектив исследователей — на основе изотопного анализа ископаемых костей эму и вомбатов (Miller et al., 2005). Предки австралийских аборигенов пришли на засушливый материк с обильно поливаемых дождями островов Малайского архипелага, где поджечь лес весьма непросто. Вероятно, переселенцы были приятно удивлены, обнаружив необыкновенно удобный и эффективный способ охоты, позволявший добыть сразу огромное количество уже поджаренной дичи. Жаль только, что в те времена люди еще не догадывались об исчерпаемости природных богатств и о том, что их деятельность может что-то нарушить в вековом природном порядке (С другой стороны, некоторые находки указывают на то, что между появлением в Австралии первых людей и вымиранием крупных сумчатых, возможно прошло довольно длительное время. Это несколько подмывает теорию молниеносного истребления мегафауны древними австралийцами). Впрочем, в иных регионах мира и по сей день можно наблюдать, как население с маниакальным упорством поджигает по весне сухую траву, невзирая на вполне реальную опасность распространения пожара на их собственные жилища и вне всякой связи с охотой. Видно, есть что-то завораживающее в зрелище горящей Земли. Покорение ЕвропыПримерно в то же время, когда сапиенсы из Юго-Восточной Азии заселили Австралию, или немного позже, их сородичи из Юго-Западной Азии начали мигрировать в северо-западном направлении и вторглись в неандертальскую Европу. Разобраться в хронологии последовавших за этим событий стало возможно благодаря последним достижениям в развитии метода радиоуглеродного датирования. Точность этого метода в последние годы возросла благодаря двум обстоятельствам. Во-первых, появились способы высококачественной очистки органических веществ, в первую очередь коллагена, выделяемого из древних костей, от всех посторонних примесей. Когда речь идет об очень древних образцах, даже ничтожная примесь постороннего углерода может привести к серьезным искажениям. Например, если в образце возрастом 40 тыс. лет окажется всего-навсего 1 % современного углерода, это приведет к уменьшению "радиоуглеродного возраста" на целых 7 тыс. лет. Как выяснилось, в большинстве древних археологических находок присутствуют такие примеси, поэтому их возраст систематически занижался. Второй источник ошибок, который наконец удалось устранить, связан с тем, что содержание радиоактивного изотопа 14С в атмосфере (а следовательно, и в органическом веществе, образовавшемся в различные эпохи) не является постоянным. Кости людей и животных, живших в периоды повышенного содержания 14С в атмосфере, изначально содержали больше этого изотопа, чем предполагалось, и поэтому их возраст опять-таки занижался. В последние годы удалось провести ряд чрезвычайно точных измерений, позволивших реконструировать колебания 14С в атмосфере в течение последних 50 тысячелетий. Для этого были использованы уникальные морские отложения в некоторых районах Мирового океана, где осадки накапливались очень быстро, гренландские льды, пещерные сталагмиты и коралловые рифы. Во всех этих случаях можно было для каждого слоя сопоставить радиоуглеродные даты с другими, полученными на основе соотношения изотопов кислорода 180/160 или урана и тория. В результате были разработаны поправочные шкалы и таблицы, позволившие резко повысить точность радиоуглеродного датирования образцов старше 25 тыс. лет. О чем же рассказали уточненные даты? Ранее считалось, что люди современного типа впервые появились на юго-востоке Европе примерно 45 тыс. лет назад. Отсюда они постепенно расселялись в западном и северо-западном направлении. Заселение Центральной и Западной Европы продолжалось, согласно "неисправленным" радиоуглеродным датировкам, примерно 7 тыс. лет (43—36 тыс. лет назад); средняя скорость продвижения — 300 м в год. Уточненные датировки показывают, что заселение началось раньше — 46 тыс. лет назад — и происходило быстрее: до 41 тыс. лет назад; скорость продвижения — до 400 м в год. Примерно с такой же скоростью позже распространялась по Европе земледельческая культура (10—6 тыс. лет назад), тоже пришедшая с Ближнего Востока. Любопытно, что обе волны расселения шли двумя параллельными путями: первый — вдоль Средиземноморского побережья от Израиля до Испании, второй — по долине Дуная, с Балкан — в Южную Германию и далее до Западной Франции. Кроме того, выяснилось, что период совместного проживания современных людей и неандертальцев в большинстве районов Европы был существенно короче, чем считалось: не 10 тыс. лет, а лишь около 6 тыс., а в некоторых районах, например на западе Франции, и того меньше — всего 1-2 тыс. лет. После этого уцелевшие неандертальцы доживали свой век в уединенных уголках Европы (таких как полуостров Гибралтар, Балканы и Крым) — вплоть до окончательного вымирания примерно 28 тыс. лет назад. По уточненным датировкам, некоторые ярчайшие образцы пещерной живописи оказались намного древнее, чем считалось. Начало эпохи ориньяк, ознаменованное появлением разнообразных сложных изделий из кости и рога (пресловутая "верхнепалеолитическая революция"), тоже сдвинулось в глубь времен — до 41 тыс. лет назад (Mellars, 2006). Все это показывает, что коренное неандертальское население Европы пало под натиском ближневосточных пришельцев значительно быстрее, чем считалось. Наверное, превосходство сапиенсов — технологическое или социальное — все-таки было слишком велико, и ни физическая сила неандертальцев, ни их выносливость, ни приспособленность к холодному климату не могли спасти обреченную расу. Как мы знаем из предыдущих глав, вопрос о том, в чем конкретно состояло это преимущество, на сегодняшний день представляется практически неразрешимым. Сапиенсы могли превосходить неандертальцев по размеру и сплоченности групп, по умению координировать действия нескольких кланов; у них могли быть более совершенные методы охоты, что привело к вытеснению неандертальцев как менее эффективных конкурентов. Некоторые данные указывают на то, что дело могло быть не только в культурно-социальных различиях, но и в биологии, в генетически обусловленных особенностях мышления (см. главу "Другое человечество"). Недавно появилось первое археологическое подтверждение гипотезы о том, что между сапиенсами и неандертальцами были прямые столкновения. В пещере Ле-Руа на юго-западе Франции среди множества типичных кроманьонских (ориньякских) артефактов найдена нижняя челюсть неандертальского ребенка с царапинами от каменных орудий. Вероятно, сапиенсы просто-напросто съели юного неандертальца, используя каменные инструменты для соскребания мяса с костей (Ramirez et al., 2009) (Л.Б. Вишняцкий (2010) указывает на спорность этой интерпретации: неандертальские признаки у челюсти не вполне очевидны; следы каннибализма можно при желании интерпретировать иначе — например, как попытки извлечь зубы для изготовления украшений; нижняя челюсть — не самая питательная часть человеческого тела. С другой стороны, похожие царапины на нижних челюстях неандертальцев из Эль-Сидрона (см. главу "Другое человечество") сомнений не вызывают, потому что на других костях остались такие же отметины: это явно следы каннибальского пиршества. Скорее всего, сапиенсы все-таки съели ребенка в пещере Лe-Pya). Прошли те времена, когда сапиенсы могли смотреть на неандертальцев как на равных и скрещиваться с ними, как в начале эпохи Великого расселения (см. выше). В ходе покорения Европы сапиенсы с неандертальцами уже не скрещивались — об этом свидетельствуют результаты прочтения неандертальского генома. Теперь бывшие родичи годились только на обед (Впрочем, некоторые антропологические данные все-таки указывают на возможную гибридизацию в Европе: по мнению С. В. Дробышевского, у поздних европейских неандертальцев часто встречаются сапиентные признаки. Может быть, скрещивание и имело место, но гибриды отличались пониженной жизнеспособностью или плодовитостью, и их потомки не дожили до наших дней. Поэтому геномы современных европейцев сохранили не больше неандертальских генов, чем геномы китайцев). Демографический факторМногие антропологи предполагают, что в начале верхнепалеолитической эпохи (примерно 45—30 тыс. лет назад), а возможно, и раньше — в среднем палеолите, популяции Homo sapiens испытали значительный демографический рост. Не исключено, что демографические факторы способствовали быстрому вытеснению сапиенсами неандертальцев и других "поздних архаичных людей" (поздние архаичные люди — условное обобщенное наименование среднепалеолитических людей, не имеющих характерных сапиентных признаков; сюда относятся не только неандертальцы, но и другие люди, которых нельзя с уверенностью отнести ни к сапиенсам, ни к неандертальцам). В этом утверждении, впрочем, есть элемент тавтологичности: если одна популяция вытесняет другую, это всегда связано с демографией (относительная, а скорее всего и абсолютная численность одной из конкурирующих популяций растет, другой — снижается). Более того, из этого следует (по определению), что первая популяция является более приспособленной, чем вторая. При этом демографический рост сам по себе является интересным объектом исследования. Он может происходить за счет разных механизмов: например, за счет снижения детской или взрослой смертности или за счет роста рождаемости. Механизмы демографического роста сапиенсов в начале верхнего палеолита точно не известны, но логично предположить, что одной из причин могло быть снижение взрослой смертности и увеличение ожидаемой продолжительности жизни взрослых людей. На такую возможность указывает тот факт, что кости и зубы неандертальцев несут в среднем больше следов болезней, травм и других признаков тяжелых жизненных условий по сравнению с костями ранних сапиенсов. Для проверки предположения о снижении взрослой смертности у средне- и верхнепалеолитических сапиенсов антрополог Эрик Тринкаус из Университета Вашингтона в Сент-Луисе проанализировал возрастной состав древних людей, чьи костные остатки позволяют определить индивидуальный возраст (Trinkaus, 2011). Для определения возраста палеоантропологи используют несколько признаков, главным из которых является степень стертости эмали коренных зубов. Учитывались только взрослые индивиды (начиная от 20 лет). Автору удалось получить достоверные данные по 59 поздним архаичным людям (преимущественно неандертальцам), 13 среднепалеолитическим сапиенсам из Азии и Северо-Восточной Африки (десять из них происходят из знаменитых пещер Схул и Кафзех на севере Израиля) и 49 сапиенсам, жившим в начале верхнего палеолита (45—25 тыс. лет назад). В каждой из трех выборок было подсчитано соотношение молодых (до 40 лет) и "старых" (40 лет и более) индивидов. Оказалось, что это соотношение практически одинаково у поздних архаичных людей и ранних сапиенсов. Из 59 "поздних архаичных людей" лишь 14 (23,7 %) дожили до сорока, из 13 среднепалеолитических сапиенсов — только один (7,7 %), из 49 верхнепалеолитических сапиенсов — 13 (26,5 %). Достоверных различий между ранними сапиенсами и неандертальцами по данному показателю не обнаружено. При этом все три выборки резко отличаются по возрастному составу от аналогичных археологических выборок, относящихся к голоцену, то есть к последним десяти тысячелетиям.
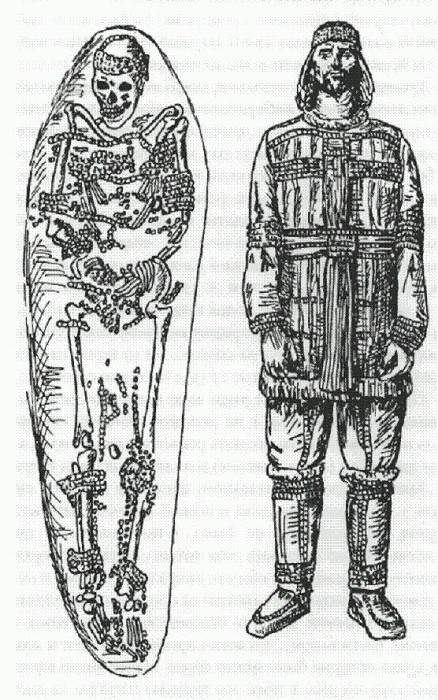
Мужчина из захоронения Сунгирь (Россия, Владимирская область, около 25 тыс. лет назад) — один из немногих палеолитических людей, которому довелось пережить свое сорокалетие. В голоценовых археологических выборках доля людей от сорока и старше составляет в среднем 39,3 %. Еще выше этот показатель у современных диких народов, не имеющих медицины (65 % двадцатилетних доживают до сорока). Тринкаус показал, что малое число людей старшего возраста в палеолитических выборках нельзя объяснить тем, что в палеолите люди по каким-то причинам предпочитали хоронить молодых, умерших в расцвете сил, а престарелых чаще оставляли без погребения (вероятность того, что скелет сохранится и попадет в руки археологов, резко возрастает, если человека похоронить). Имеющиеся данные указывают как раз на обратную картину: среди палеолитических людей, которые были сознательно похоронены своими соплеменниками, доля пожилых существенно выше, чем в среднем по всей выборке. Из 59 поздних архаичных людей похоронено было 18, из них 44,4 % старше сорока; из 13 среднепалеолитических сапиенсов похоронено 11 (9,1 % старше сорока); из 49 верхнепалеолитических сапиенсов похоронено 27 (33,3 % старше сорока). По-видимому, древние люди чаще предавали земле как раз пожилых соплеменников, а не молодых. Поэтому избирательность погребений могла исказить реальную картину лишь в сторону завышения (а не занижения) доли пожилых среди умерших. Таким образом, радикального снижения взрослой смертности у сапиенсов в среднем и первой половине верхнего палеолита, по-видимому, не было. Оно произошло позже, но когда именно — сказать пока трудно. Средне- и верхнепалеолитические сапиенсы обычно умирали молодыми, и по этому демографическому показателю их популяции не отличались от неандертальских. Во всех человеческих коллективах того времени, по-видимому, численно преобладали дети и подростки, среди которых было много сирот. Малое число взрослых людей в группе вело к тому, что трудовая нагрузка на каждого из них была очень велика, что вряд ли способствовало их долгой жизни. К тому же и дедушек с бабушками, которые могли бы помочь нянчиться с малышней, в группах почти не было. Чем же был обусловлен предполагаемый рост численности сапиенсов в верхнем палеолите, если взрослая смертность не уменьшилась? Одно из двух: либо выросла плодовитость (женщины стали чаще рожать), либо снизилась детская и подростковая смертность. Проверить это крайне трудно, потому что детей в те времена хоронили редко, и их останки имели мало шансов сохраниться. Венера и флейтаСлучайно ли, нет ли, но заселение сапиенсами Европы и вытеснение коренного неандертальского населения совпадает в пространстве и времени с началом ориньякской эпохи, ознаменованной новыми технологиями изготовления орудий (верхнепалеолитические технологии обработки камня и кости настолько сложны и изощренны, что находятся далеко за пределами возможностей не только простых смертных, но и большинства археологов), а также внезапным появлением и бурным расцветом искусства. Искусство эпохи ориньяк — это вам не скромные ожерелья из ракушек и геометрические царапины на скорлупе страусиных яиц, как в культурах стилбей и ховьесонс пурт в южной Африке 70—60 тыс лет назад (см. главу "От эректусов к сапиенсам"). На этот раз наши предки создали нечто большее: настоящую живопись, музыку, скульптуру. Причем произошло все это довольно быстро. За пределами Европы искусство появляется, по-видимому, на несколько тысячелетий позже. О древнейшей в мире скульптуре мир узнал совсем недавно — в 2009 году. Нашли ее в пещере Холе-Фельс ("Полая скала") на юго-западе Германии. Пещерные отложения в Холе-Фельс подразделяются на 18 слоев. Нижние четыре слоя относятся к среднему палеолиту и содержат следы пребывания неандертальцев. Над ними расположены отложения верхнего палеолита, когда в пещере жили люди современного типа. Из этих верхнепалеолитических слоев шесть нижних относятся к эпохе ориньяк — древнейшему этапу европейского верхнего палеолита, над ними залегают слои граветтской и мадленской эпох. Сенсационная находка, о которой сообщил археолог Николас Конард из Тюбингенского университета, — вырезанная из мамонтовой кости женская статуэтка, происходящая из самого нижнего слоя ориньякской эпохи (Conard, 2009). Непосредственно под этим слоем идут уже среднепалеолитические неандертальские слои, отделенные от верхнепалеолитических отложений тонким прослоем "стерильной" глины. Этот прослой очень важен: он означает, что сапиенсы никого не выгоняли, а пришли в давно опустевшую пещеру. Что случилось с неандертальцами из Холе-Фельс? Никто не знает. В 2003 г. Конард нашел двумя слоями выше — в третьем снизу ориньякском слое — три костяные фигурки: лошадиную голову, водоплавающую птицу и териантропа (странное существо с признаками человека и кошки). Они считались одними из самых древних, — а возможно, и самыми древними произведениями искусства, однако новая находка еще древнее. Точный возраст статуэтки и вмещающих отложений попытались определить радиоуглеродным методом. Анализировались в основном фрагменты костей животных, найденные в непосредственной близости от статуэтки. В этих костях сохранилось довольно много коллагена, который и использовался для анализов, проводившихся независимо в нескольких лабораториях. Однако результаты получились не совсем однозначные. "Исправленные" (откалиброванные) радиоуглеродные датировки варьируют примерно от 35—36 до 44 тыс. лет назад. Такой разброс может быть связан со множеством причин — например, со случайным загрязнением проб посторонними примесями. По мнению автора, наиболее вероятный возраст статуэтки — около 40 тыс. лет, что соответствует самому началу эпохи ориньяк. Этот вывод основан на том, что находка сделана в самой нижней части метровой толщи ориньякских отложений. При этом нужно иметь в виду, что пещера Холе-Фельс — чуть ли не самая большая и удобная в этом районе и к тому же хорошо заметна, поэтому пришедшие сюда сапиенсы должны были заселить ее одной из первых. Статуэтка была расколота на шесть кусков, которые все были найдены в пределах одного квадрата 25 х 25 см. У склеенной скульптуры недостает левой руки и плеча, но Конард надеется, что они со временем найдутся — ведь находка была сделана на самом краю раскопа, рядом с его вертикальной глиняной стенкой, в толще которой, возможно, и скрываются недостающие части древнейшей скульптуры. Статуэтка довольно реалистична с точки зрения анатомии, но кое в чем древний скульптор дал волю фантазии — иначе какое же это искусство? Гипертрофированные женские половые признаки роднят ее со знаменитыми "палеолитическими венера- ми" более поздней граветтской эпохи (начавшейся 27—28 тыс лет назад). Еще одной интересной особенностью статуэтки является полное отсутствие головы. Видимо, древний скульптор считал эту часть женского тела наименее существенной. Вместо головы он вырезал аккуратное колечко, чтобы можно было носить фигурку на шнурке. Колечко слегка отполировано изнутри: вероятно, кто-то ее действительно носил — может быть, в качестве амулета. Талия Холе-фельсской Венеры лишь ненамного уже широких плеч и бедер. Живот разукрашен горизонтальными линиями, которые, вероятно, символизируют какое-то одеяние. Линии были процарапаны острыми каменными орудиями, причем древний скульптор многократно проводил острием по одному и тому же месту и прикладывал значительные усилия, чтобы борозда в твердой мамонтовой кости стала достаточно глубокой. В отличие от типичных "палеолитических венер" грудь у Холе-фельсской Венеры не висит, плечи очень широкие, а живот почти плоский. Это не просто древнее произведение искусства. Это еще и самое старое дошедшее до нас изображение человека. Уникальное окно в прошлое, единственный способ своими глазами взглянуть на человека раннеориньякского времени. Жаль, что оно все-таки слишком стилизованное и без головы.
«Палеолитические венеры» Новая находка заставила пересмотреть сложившиеся представления о зарождении палеолитического искусства. До сих пор считалось, что для ориньякской эпохи были характерны только изображения животных, в основном крупных и агрессивных, а также объектов, связанных с шаманскими культами (вроде вышеупомянутого человека-кошки). Считалось, что женское тело стало источником вдохновения для палеолитических скульпторов лишь в следующую, граветтскую эпоху. Теперь очевидно, что это творческое направление присутствовало в верхнепалеолитической художественной культуре с самого ее зарождения. Скорее всего, женские фигурки с преувеличенными половыми признаками прямо или косвенно символизировали плодородие (Медникова, 2007). Возможно, тот же смысл несли и многочисленные схематические изображения мужских и женских половых органов, выцарапанные на камнях, и фаллические скульптуры возрастом 35—36 тыс. лет, найденные на юго-западе Франции. Вскоре после публикации статьи о древнейшей Венере германские археологи сообщили еще об одной сенсационной находке, сделанной ими в той же пещере и в том же самом слое, возраст которого составляет около 40 тыс. лет (Conard et al., 2009). Флейта, вырезанная из лучевой кости белоголового сипа, была найдена 17 сентября 2008 года. Белоголовый сип — крупная птица (размах крыльев 230-265 см), кости которой идеально подходят для изготовления флейт. Длина сохранившейся части флейты — 21,8 см, диаметр — 8 мм. У флейты пять отверстий для пальцев и два глубоких V-образных выреза на том конце, в который дул музыкант. Противоположный конец инструмента (длиной в несколько сантиметров) не сохранился. Исследователи предполагают, что из инструмента можно было извлечь не менее пяти нот. В том же слое найдены фрагменты двух других флейт, выточенных из мамонтового бивня. Над этими инструментами древним мастерам пришлось потрудиться побольше, чем над дудочкой из полой птичьей кости. Сначала вырезалась заготовка, которая затем разделялась вдоль на две половинки. В каждой их половинок вытачивали желобок, а затем половинки каким-то образом склеивали. Ранее в ориньякских отложениях юго-западной Германии уже было найдено несколько обломков флейт из птичьих костей и мамонтового бивня. В общей сложности, вместе с новыми находками, описаны фрагменты четырех "птичьих" и четырех "мамонтовых" флейт. Ценность новых находок прежде всего в том, что они имеют более точную стратиграфическую привязку: о них можно с уверенностью сказать, что они были изготовлены в самом начале ориньякской эпохи, то есть вскоре после появления в Европе людей современного типа. Кроме того, находки показали, что уже в самом начале верхнего палеолита — сразу после "верхнепалеолитической революции" — игра на музыкальных инструментах была обычной, широко распространенной практикой у наших предков. Возможно, музыка способствовала сплочению коллективов кроманьонцев и в какой-то мере помогала их быстрой экспансии и вытеснению коренного европейского населения. Ранее в Словении была найдена медвежья кость возрастом 43 тыс. лет с двумя круглыми дырочками, которая была интерпретирована как неандертальская флейта. Но в настоящее время большинство специалистов считают эту интерпретацию ошибочной. По-видимому, дырочки были проделаны зубами хищника. Все бесспорные палеолитические флейты, найденные за пределами юго-западной Германии, моложе 30 тыс. лет. Флейта из Холе-Фельс найдена всего в 70 см от древнейшей женской статуэтки, и археологи не исключают, что эти предметы были каким-то образом связаны друг с другом. Восточная Европа заселялась в то же время, что и Западная Хронология заселения сапиенсами Восточной Европы изучена хуже, чем Западной, и дело тут не столько в недостатке изученных стоянок первобытного человека, сколько в том, что их очень трудно точно датировать. Уникальный комплекс верхнепалеолитических стоянок Костёнки находится в Воронежской области на западном берегу Дона. Здесь на небольшой территории сконцентрировано около 30 стоянок разного возраста, причем некоторые из них — многослойные, то есть люди обитали здесь в течение длительного времени. Найдены жилища из мамонтовых костей, "палеолитические венеры", фрагменты украшений и другие признаки высокой культуры. Человеческих костей в самых нижних из этих слоев, к сожалению, найти не удалось (за исключением отдельных зубов), зато там много каменных и костяных изделий, характерных для верхнего палеолита. В 2007 году археолог М. В. Аникович из Института истории материальной культуры (Санкт-Петербург) и его коллеги из разных научных учреждений России, США, Великобритании и Италии опубликовали в журнале Science статью, в которой подвели итоги многолетних усилий по установлению точного возраста этих стоянок (Anikovich et al., 2007). Датировать находки археологам помог прослой вулканического пепла возрастом около 40 тыс. лет, который прослеживается в ряде районов Центральной и Восточной Европы. Возраст пепла определен при помощи радиоизотопного аргон-аргонового метода. Самые древние слои, содержащие верхнепалеолитические изделия, в Костёнках находятся ниже этого прослоя — стало быть, они старше. Возраст определяли также при помощи радиоуглеродного, люминесцентного и палеомагнитного методов и споро-пыльцевого анализа. Вся совокупность полученных данных свидетельствует о том, что сапиенсы — носители верхнепалеолитической культуры — жили на территории Воронежской области уже 42—45 тыс. лет назад (По самым последним данным, древнейший верхний палеолит в Костёнках датирован 52 тыс. лет назад (С. В. Дробышевский, личное сообщение)). Набор артефактов, найденный в самых древних слоях, несомненно является верхнепалеолитическим. Об этом свидетельствуют, в частности, просверленные раковины моллюсков — остатки древних подвесок и ожерелий, а также разнообразные изделия из кости, включая вырезанный из мамонтового бивня округлый объект, возможно, являющийся незаконченным скульптурным изображением человеческой головы. Однако этот комплекс все-таки сильно отличается от типичной ориньякской культуры, характерной для Центральной и Западной Европы. Возможно, сапиенсы, впервые объявившиеся на Балканах и в Восточной Европе (в долине Дона) примерно в одно и то же время, были представителями разных племен и пришли из разных мест: первые, скорее всего, из Леванта и Малой Азии, а вторые — из Закавказья или Центральной Азии. Проблема перехода от среднего к верхнему палеолиту — одна из самых спорных в археологии. В Западной и Центральной Европе этот переход достаточно резкий, что соответствует быстрому вытеснению неандертальцев с их среднепалеолитической (мустьерской) культурой сапиенсами с их верхнепалеолитическими (ориньякскими) технологиями. В Центральной Азии, как мы уже упоминали в главе "Другое человечество", наблюдается более плавный переход от среднего к верхнему палеолиту с явными признаками культурной преемственности. В Восточной Европе переход происходил скорее по западноевропейскому типу, то есть был резким, скачкообразным. В районе Костёнок имеются и стоянки, которые можно трактовать как среднепалеолитические или переходные от среднего к верхнему палеолиту (так называемая стрелецкая культура). Человеческих костей там тоже не нашли, но предполагается, что эти стоянки принадлежали местным неандертальцам. Стрелецкие комплексы артефактов встречаются и ниже, и выше прослоя вулканического пепла, то есть предполагаемые неандертальцы, по-видимому, какое-то время жили бок о бок с пришельцами-сапиенсами. При этом стрелецкие стоянки резко отличаются от верхнепалеолитических. Там отсутствуют изделия из кости и рога, там нет украшений и произведений искусства. Два человечества жили рядом, но ничего не хотели друг о друге знать! Сапиенсам с их высокой культурой неандертальцы должны были казаться "грязными дикарями" или вообще "животными". Об этом мы можем судить по аналогичным ситуациям в недавнем историческом прошлом. Что думали неандертальцы, мы, к сожалению, вряд ли когда-нибудь узнаем. Кроме того, некоторые каменные изделия, обнаруженные на древнейших верхнепалеолитических стоянках в Костёнках, изготовлены из камня, который можно было найти лишь в 100—150 км от стоянки, а ракушки, найденные там, могли быть принесены только с черноморских берегов, расположенных на расстоянии 500 км. На стрелецких стоянках все изделия изготовлены из местных материалов. Наконец, существует еще одно важное различие между среднепалеолитическими и одновозрастными верхнепалеолитическими стоянками этого района. На первых встречаются в основном кости крупных животных (таких как лошадь и северный олень). На вторых кости крупных животных редки, а преобладают кости зверей мелкого и среднего размера (заяц, лиса, песец, волк), а также птиц. Все это говорит о том, что люди современного типа пришли в Восточную Европу около 45—42 тыс. лет назад, уже обладая высокоразвитой верхнепалеолитической культурой, и постепенно вытеснили обитавшее здесь древнее население (предположительно неандертальцев). Назад в АфрикуВнеафриканское человечество, по-видимому, происходит в основном от группы сапиенсов, вышедших из Африки через Баб-эль-Мандебский пролив. Эти люди были носителями митохондриальных линий М и N. Что касается африканцев (особенно тех, что живут к югу от Сахары), то они, как считалось до недавних пор, в основном являются потомками тех сапиенсов, которые никогда не покидали своей прародины. Это подтверждается гораздо более высоким разнообразием типов мтДНК у африканцев по сравнению с не-африканцами (у первых сохранились разнообразные древние варианты мтДНК, а у не-африканцев — только вариации типов М и N). Недавно к этой картине была добавлена новая важная деталь. Выяснилось, что у коренных жителей некоторых северных и восточных районов Африки часто встречаются два нетипичных для африканцев варианта мтДНК. Один из этих вариантов (M1) относится к группе М, которая распространена в Южной и Юго-Восточной Азии и в Океании. Второй вариант (U6) относится к распространенной по всей Евразии подгруппе R группы N (Olivieri et al., 2006) Проще всего было бы предположить, что это результат каких-то сравнительно поздних миграций азиатов в Африку. Но это предположение не подтвердилось. Ученые определили последовательность нуклеотидов мтДНК у 51 носителя варианта M1 и 30 носителей варианта U6. Сравнение этих последовательностей с другими известными последовательностями человеческой мтДНК показало, что обе линии (M1 и U6) — весьма древние. Они, несомненно, происходят от евразийских групп М и N, но обособились от них давно — около 40 тыс. лет назад. Крайне любопытно, что ближайшей родственницей линии U6 является линия U5, которая, по мнению генетиков, появилась около 50 тыс. лет назад и могла встречаться у носителей культуры ориньяк, то есть людей современного типа, заселивших неандертальскую Европу и достигших небывалого культурного расцвета (хотя прямых палеогенетических подтверждений эта идея пока не имеет). Приняв во внимание множество дополнительных фактов, исследователи предположили следующее. Вышедшие из Африки люди, вероятно, долгое время были отрезаны от своей прародины и от Европы пустыней, которая простиралась от Северной Африки до Центральной Азии. Около 46 тыс. лет назад климат улучшился, и люди проникли с востока в Левант. Это были носители вариантов мтДНК U5, U6 и M1. По-видимому, где-то здесь, в Западной Азии, они приобщились к верхнепалеолитической культуре — самым передовым технологиям того времени. Может быть, они сами ее и создали. Из Леванта они стали расселяться дальше: одни в Северную Африку (где сохранились варианты U6 и M1), другие — в Европу, где сохранился вариант U5. Из этого следует, что древнейшие африканские и европейские верхнепалеолитические культуры — даббан и ориньяк, — возможно, происходят от общего западноазиатского корня. Открытие АмерикиСледя за приключениями наших предков, мы постепенно переходили от "еще обезьян" к "уже не совсем обезьянам", "то ли обезьянам, то ли людям", "почти людям" и наконец добрались до "совсем уж точно людей". Мы увидели, что человеческие черты развивались понемногу, не все сразу, что они не образуют единого комплекса неразрывно связанных друг с другом признаков. Постепенно на этом пути снижалась роль биологических факторов (генов, мутаций, отбора) и на первый план выходили факторы культурные — накопление знаний и традиций, передающихся из поколение в поколение негенетическим путем. Разумеется, в зачаточной форме культурная эволюция существовала уже у древнейших гоминид — как существует она и сегодня у шимпанзе. Разумеется, биологическая эволюция не прекратилась и в современном человечестве (об этом мы подробнее поговорим в "Заключении"). Провести четкую грань, найти точку во времени, когда биологическая эволюция гоминид уступила место культурной, невозможно. Точно так же, как невозможно точно указать момент, когда "нечеловеческая обезьяна" превратилась в "человеческую". Хотите — считайте, что это произошло с началом изготовления каменных орудий, или с началом роста мозга у хабилисов, или с быстрым ростом мозга у эректусов, или с переходом от олдувая к ашелю, или с освоением огня, или с появлением гейдельбергских людей с таким же большим мозгом, как у нас, или с появлением "анатомически современных" сапиенсов, или с появлением ожерелий на юге Африки, или с "верхнепалеолитической революцией". Дело это чисто условное и, по правде сказать, довольно бессмысленное: все равно что пытаться абсолютно точно установить сумму, начиная с которой мало денег" превращается в "много денег". Однако нам постоянно приходится принимать именно такие условные и научно не обоснованные решения, ведь наши действия поневоле дискретны. При встрече с соперником животные демонстрируют так называемую реакцию борьбы или бегства (fight or flight response) — либо вступают в схватку, либо убегают. Нельзя наполовину броситься в бой, а наполовину удрать. Из-за необходимости постоянно принимать дискретные решения наше мышление работает "категориально": мы то и дело пытаемся найти четкие границы между понятиями, в том числе и там, где их на самом деле нет, и разделить непрерывный ряд (континуум) на отрезки, приклеив к ним слова-ярлыки (см. главу "Душевная механика", кн. 2). Сейчас нам нужно методом волевого решения выбрать момент, на котором рассказ об эволюции гоминид можно прекратить — а о том, что было дальше, пусть в других книгах рассказывают историки с социологами. По-моему, вступление наших предков в эпоху верхнего палеолита — вполне подходящий момент для проведения такой условной грани. Но об одном более позднем событии — заселении Северной и Южной Америки — все-таки следует рассказать. Уж очень масштабная была акция. Никаких других гоминид, кроме сапиенсов, в Новом Свете никогда не было. Все сообщения о находках такого рода впоследствии были опровергнуты. Первые люди — верхнепалеолитические охотники-сапиенсы — проникли в Америку из Азии, воспользовавшись сухопутным мостом — Берингией, которая во времена оледенений соединяла Чукотку с Аляской. Согласно одной из гипотез, примерно 13,5 тыс. лет назад переселенцы впервые прошли по узкому коридору между ледниками в западной Канаде и очень быстро — всего за несколько веков — расселились по всему Новому Свету вплоть до южной оконечности Южной Америки. Вскоре они изобрели крайне эффективное охотничье оружие — дротики с плоскими, симметричными и устрашающе острыми кремневыми наконечниками (так называемая культура кловис) — и перебили большую часть мегафауны (крупных животных) на обоих материках. Однако новые факты, полученные генетиками и археологами, показывают, что в действительности история заселения Америки была несколько более сложной. Специалисты продолжают энергично спорить о ее деталях. Критический обзор различных версий дан в книге Ю. В. Березкина "Мифы заселяют Америку" (2007). Мы же будем в основном придерживаться фактов и гипотез, изложенных в большой обзорной статье, опубликованной недавно в журнале Science (Goebel et al., 2008). ДАННЫЕ ГЕНЕТИКИ. Азиатское происхождение коренных американцев не вызывает сомнений. В Америке распространены пять вариантов митохондриальной ДНК (А, В, С, D, X), причем все они характерны также для коренного населения Южной Сибири от Алтая до Амура (Захаров, 2003). Митохондриальная ДНК, извлеченная из костей древних американцев, также имеет явно азиатское происхождение. Попытки установить на основе анализа мтДНК и Y-xpoмосомы время разделения азиатских и американских популяций пока дают довольно противоречивые результаты (получающиеся датировки варьируют от 25 до 15 тыс. лет). Несколько более надежными считаются оценки времени начала расселения палеоиндейцев к югу от ледникового щита: 16,6—11,2 тыс. лет. Эти оценки основаны на анализе трех вариантов подгруппы C1, широко распространенных среди индейцев, но не вcтречающихся в Азии. По всей видимости, эти варианты мтДНК возникли уже в Новом Свете. Более того, анализ географического распространения различных вариантов мтДНК среди современных индейцев показал, что наблюдаемую картину легче объяснить исходя из допущения, что расселение началось ближе к началу, а не к концу указанного временного интервала (то есть скорее 15—16, а не и—12 тыс. лет назад).
Гигантский броненосец глиптодон — один из ярких представителей американской мегафауны, вымерший около 10 тыс. лет назад. Палеоиндейцы иногда хоронили своих сородичей в панцирях глиптодонов, как в саркофагах. Некоторые антропологи высказывали предположение о "двух древних волнах" заселения Америки (в рамках этой теории более поздний приход в Америку предков эскимосов рассматривается как третья волна). Идея "двух волн" основывалась на том, что древнейшие человеческие черепа, найденные в Новом Свете, заметно отличаются по ряду размерных показателей от черепов современных индейцев. Но генетические данные не подтверждают идею двух волн. Наблюдаемое распределение генетических вариаций свидетельствует о том, что все генетическое разнообразие коренных американцев происходит из единого предкового азиатского генофонда и широкое расселение людей по обеим Америкам имело место только единожды. Так, во всех изученных популяциях индейцев от Аляски до Бразилии встречается один и тот же аллель (вариант) одного из ядерных микросателлитных локусов (микросателлитные локусы — участки генома, состоящие из многократно повторяющихся коротких последовательностей нуклеотидов. Микросателлиты очень полиморфны, то есть различаются у разных особей одного и того же вида (например, по числу повторов). Поэтому их часто используют в качестве маркёров для оценки генетического родства и выяснения генеалогий), который не встречается нигде за пределами Нового Света, за исключением чукчей и коряков. Это говорит о том, что все индейцы произошли от единой предковой популяции, а не от нескольких разных. У древнейших американцев, судя по данным палеогенетики, были распространены те же группы мтДНК, что и у современных индейцев. ДАННЫЕ АРХЕОЛОГИИ. Уже 32 тыс. лет назад люди — носители верхнепалеолитической культуры — заселили Северо- Восточную Азию вплоть до побережья Северного Ледовитого океана. Об этом свидетельствуют, в частности, археологические находки, сделанные в низовьях реки Яны, где обнаружены изделия из мамонтовой кости и рогов шерстистого носорога (среди сибирских археологов имеются разногласия по поводу датировки заселения северо-востока Азии). Заселение Арктики, по-видимому, произошло в период сравнительно теплого климата перед наступлением последнего ледникового максимума. Не исключено, что уже в эту отдаленную эпоху обитатели азиатского северо-востока проникали на Аляску. Там найдено несколько костей мамонта возрастом около 28 тыс. лет, возможно, подвергавшихся обработке. Однако искусственное происхождение этих объектов спорно, а никаких каменных орудий или других явных признаков присутствия человека поблизости не обнаружено. Древнейшие бесспорные следы присутствия человека на Аляске — каменные орудия, очень похожие на те, что производились верхнепалеолитическим населением Сибири, — имеют возраст 14 тыс. лет. Дальнейшая археологическая история Аляски довольно сложна. Здесь обнаружено много стоянок возрастом 12—13 тыс. лет с разными типами каменной индустрии. Возможно, это свидетельствует об адаптации местного населения к быстро меняющемуся климату, но может отражать и миграции племен. 40 тыс. лет назад большая часть Северной Америки покрылась ледниковым щитом, который перегородил путь с Аляски на юг. Сама Аляска льдом не покрывалась. В периоды потеплений в ледниковом щите открывались два коридора — вдоль Тихоокеанского побережья и к востоку от Скалистых гор, — по которым древние жители Аляски могли пройти на юг. Коридоры были открыты 32 тыс. лет назад, когда люди появились в низовьях Яны, но 24 тыс. лет назад они снова закрылись. Люди не успели ими воспользоваться. Прибрежный коридор снова открылся около 15 тыс. лет назад, а восточный — несколько позже, 13,0—13,5 тыс. лет назад. Впрочем, древние охотники теоретически могли обойти препятствие морским путем. На островах Санта-Роза и Сан- Мигель у побережья Калифорнии обнаружены следы присутствия человека возрастом 12800—11500 лет. Это означает, что население Америки в те времена уже хорошо знало, что такое лодка или плот (в 2010 году на острове Крит были обнаружены каменные орудия возрастом не менее 130 тыс. лет. Это свидетельствует о неплохих навыках мореплавания у каких-то человеческих популяций еще в среднепалеолитическую эпоху). Детально документированная археологическая история Америки к югу от ледника начинается с культуры кловис. Расцвет этой культуры охотников на крупного зверя был стремительным и скоротечным. Согласно последним уточненным радиоуглеродным датировкам, древнейшие материальные следы культуры кловис имеют возраст 13 200—13 100 лет, а самые молодые — 12 900—12 800 лет. Культура кловис так быстро распространилась по обширным территориям Северной Америки, что археологи пока не могут определить район, в котором она впервые появилась: точность методов датирования оказывается недостаточной для этого. Всего через 2—4 столетия после своего появления культура кловис столь же стремительно исчезла. По-видимому, кловисцы были кочевыми охотниками-собирателями, способными быстро перемещаться на большие расстояния. Их каменные и костяные орудия были весьма совершенны, многофункциональны, изготавливались при помощи оригинальных методик и весьма ценились своими обладателями. Каменные орудия производились из высококачественного кремня и обсидиана — материалов, которые далеко не везде можно найти, поэтому люди берегли их и носили с собой, порой унося на сотни километров от места изготовления. Стоянки культуры кловис представляют собой небольшие временные лагеря, где люди не жили подолгу, а останавливались лишь затем, чтобы съесть очередного убитого крупного зверя. Кроме того, на юго-востоке США и в Техасе найдены громадные скопления кловисских артефактов — до 650 тыс. штук в одном месте. В основном это отходы каменной индустрии. Очевидно, здесь у кловисцев были главные "каменоломни" и "оружейные мастерские". Излюбленной добычей кловисцев были хоботные — мамонты и мастодонты. В Северной Америке обнаружено как минимум 12 бесспорных кловисских "мест забоя и разделки хоботных" (это их официальное научное название (proboscidean kill and butchery sites)). Это очень много, учитывая кратковременность существования культуры кловис. Для сравнения, во всем верхнем палеолите Евразии (что соответствует периоду времени примерно в 30 тыс. лет) обнаружено всего шесть таких стоянок. Вполне возможно, что люди кловис внесли немалый вклад в вымирание американских хоботных. Не брезговали они и более мелкой добычей — бизонами, оленями, зайцами и даже рептилиями и амфибиями.
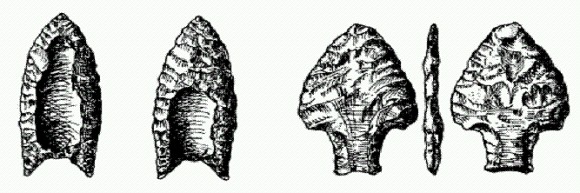
Слева — типичный наконечник культуры кловис. Справа — рыбообразный наконечник, найденный в Белизе. Культура кловис проникла в Центральную и Южную Америку, но здесь она не получила такого широкого распространения, как в Северной (найдено лишь небольшое количество типичных кловисских артефактов). Зато в Южной Америке обнаружены палеолитические стоянки с другими типами каменных орудий, в том числе с характерными рыбообразными наконечниками. Некоторые из этих южноамериканских стоянок по возрасту перекрываются с кловисскими. Раньше считалось, что культура "рыбных" наконечников произошла от кловисской, но уточнение датировок, проведенное недавно, показало, что, возможно, обе культуры происходят от некого общего и пока еще не обнаруженного предка". На одной из южноамериканских стоянок найдены кости вымершей дикой лошади. Это значит, что первопоселенцы Южной Америки, вероятно, тоже внесли свой вклад в истребление крупных животных. Жаль, не знали палеоиндейцы, на что еще годятся лошади: если бы они не съели их, а приручили, как впоследствии ламу, еще неизвестно, как повернулась бы дальнейшая история. В течение второй половины ХХ века археологи неоднократно сообщали о находках более древних следов присутствия человека в Америке, чем стоянки культуры кловис. Большинство этих находок после тщательных проверок оказывались более молодыми. Однако для нескольких стоянок "докловисский" возраст сегодня признается большинством специалистов. В Южной Америке это стоянка Монте-Верде в Чили, возраст которой — 14600 лет. В штате Висконсин, у самого края существовавшего в те времена ледникового щита, обнаружены две стоянки древних любителей мамонтятины — то ли охотников, то ли падальщиков. Возраст стоянок — от 14200 до 14800 лет. В том же районе найдены кости мамонтовых ног с царапинами от каменных орудий; возраст костей — 16 тыс. лет, правда самих орудий поблизости так и не нашли. В Пенсильвании, Флориде, Орегоне и других районах США сделано еще несколько находок, с разной степенью достоверности указывающих на присутствие людей в этих местах 14—15 тыс. лет назад. Немногочисленные находки, возраст которых был определен как еще более древний (свыше 15 тыс. лет), вызывают у специалистов большие сомнения. ПРОМЕЖУТОЧНЫЕ ИТОГИ. Хотя некоторые палеоиндейские черепа отличаются от современных, результаты генетического анализа свидетельствуют в пользу версии о происхождении всего коренного населения Америки — как древнего, так и современного — от одной и той же популяции выходцев из Северо-Восточной Азии, которые, в свою очередь, пришли туда из южной Сибири. Первые люди появились на северозападной окраине североамериканского континента не ранее 30 и не позднее 13 тыс. лет назад, скорее всего между 22 и 16 тыс. Судя по молекулярно-генетическим данным, расселение из Берингии на юг началось не ранее чем 16,6 тыс. лет назад, причем размер популяции "основателей", от которой произошло все население обеих Америк к югу от ледника, не превышал 5 тыс. человек. Теория многократных волн заселения не подтвердилась (за исключением эскимосов, алеутов и некоторых других племен, которые пришли из Азии значительно позже, но заселили только крайний север американского континента). Опровергнута также и теория об участии европейцев в древней колонизации Америки.
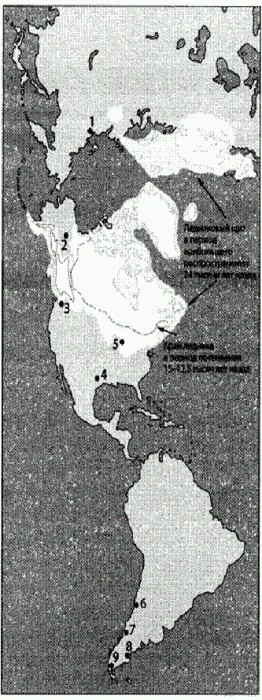
Белым цветом обозначен ледниковый щит в период наибольшего распространения 24 тыс. лет назад, пунктирной линией обведен край ледника в период потепления 15—12,5 тыс. лет назад, когда открылись два "коридора" с Аляски на юг. Черными точками показаны места археологических находок: 1 — стоянка в низовьях Яны (32 тыс. лет); 2 — кости мамонта с возможными следами обработки (28 тыс. лет); 3 — Кенневик (здесь найден хорошо сохранившийся скелет "палеоиндейца "); 4 — самая большая «мастерская» культуры кловис в Техасе (630 тыс. артефактов); 5 — древнейшие находки в штате Висконсин (14,2—14,8 тыс. лет); 6 — южноамериканская стоянка с костями лошади (13,1 тыс. лет); 7 — Монте-Верде (14,6 тыс. лет); 8,9 — здесь найдены рыбообразные наконечники, возраст которых (12,9—13,1 тыс. лет) совпадает со временем существования культуры кловис. По рисунку из Goebel et al., 2008. Одно из важных достижений последних лет состоит в том, что кловисцы, по-видимому, больше не могут считаться первопоселенцами обеих Америк к югу от ледника. Эта теория предполагает, что все более древние археологические находки должны быть признаны ошибочными, а с этим на сегодняшний день уже никак нельзя согласиться. Кроме того, эта теория не подтверждается данными по географическому распространению генетических вариаций среди индейского населения, которые свидетельствуют о более раннем и не таком стремительном заселении обеих Америк. По-видимому, обе Америки были заселены примерно 15 тыс. лет назад — практически сразу после того, как открылся прибрежный "коридор", позволивший обитателям Аляски проникнуть на юг сухим путем. Находки в Висконсине и Чили показывают, что 14,6 тыс. лет назад обе Америки уже были обитаемы. У первых американцев наверняка были лодки или хорошие плоты, что могло способствовать их быстрому расселению вдоль тихоокеанского побережья. Это подтверждается новейшими результатами изучения палеоиндейских стоянок на островах Санта-Роза и Сан-Мигель у побережья Калифорнии. Древние обитатели островов были хорошо приспособлены к жизни на океанском побережье; они охотились на морских птиц, ловили рыбу и морского зверя, собирали моллюсков (Erlandson et al., 2011). Второй предположительный путь ранних миграций — на восток вдоль южного края ледникового щита до Висконсина и далее. Вблизи ледника могло быть особенно много мамонтов, за которыми и шли древние охотники.
Неандерталец.
Barbourofelis — североамериканский представитель саблезубых кошек.
Пещерный лев Panthera leo spelaea.
Один из самых ярких и необычных представителей плейстоценовой фауны Австралии — сумчатый лев Thylacoleo.
Усть-Каракол — пещера на Алтае (недалеко от Денисовой), где, возможно, жили неандертальцы или денисовцы. Правда, человеческих костей там пока не нашли — только каменные орудия. См. главу "Другое человечество".
Верхняя часть раскопа в Денисовой пещере. Внизу видны слои 9 и 10. Полосатая толща над ними — голоценовые отложения (последние 10 тыс. лет). В это время в пещере стали регулярно жечь костры.
Нижняя часть раскопа в Денисовой пещере: от самых нижних слоев, относящихся к началу среднего палеолита, до верхнепалеолитических.
Среднепалеолитические орудия неандертальцев из грота Тешик-Таш (Узбекистан). См. главу "Другое человечество".
Неандертальский мальчик из грота Тешик-Таш (Узбекистан). Реконструкция М. М. Герасимова. См. главу "Другое человечество".
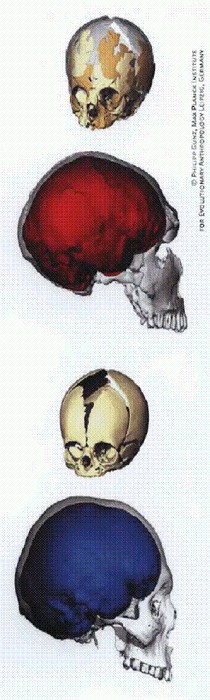
Черепа новорожденных и взрослых представителей Homo neanderthalensis и Н. sapiens.
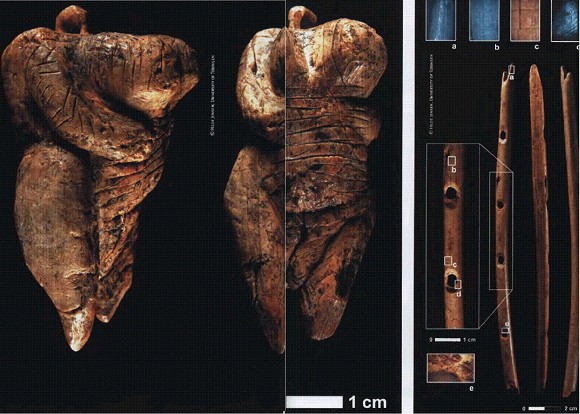
Находки из пещеры Холе-Фельс, Германия, 40 тыс. лет. Слева - так называемая Холе-фельсская Венера. Справа: флейта, найденная неподалеку от древнейшей в мире статуэтки. a-е — следы обработки птичьей кости каменными орудиями. См. главу "Великое расселение сапиенсов".
Верхнепалеолитические костяные изделия. Вверху: фигурки "палеолитических венер". См. главу "Великое расселение сапиенсов".
Верхнепалеолитические орудия европейских Homo sapiens (кроманьонцев). См. главу "Великое расселение сапиенсов".
Черепа кроманьонцев.
Мужчина Сунгирь-1 — верхнепалеолитический охотник на мамонтов. Владимирская обл., около 25 тыс. лет. Реконструкция М. М. Герасимова. См. главу "Великое расселение сапиенсов".
Пока самка придирчиво осматривает беседку, самец пытается обратить ее внимание на лучшие драгоценности в своей коллекции. У каждого вида шалашников свои представления об эстетике. Атласным шалашникам (Ptilonorhynchus violaceus) нравится синий цвет, а большие серые шалашники (P. nuchalis) любят красный, зеленый и белый, но терпеть не могут все желтое. См. главу "Происхождение человека и половой отбор".
Вверху: драка коршунов за территорию. Внизу: черные коршуны в Испании украшают свои гнезда обрывками белых пластиковых пакетов, причем количество украшений свидетельствует о здоровье и силе хозяев гнезда. Молодые коршуны почти не украшают свои гнезда (А), птицы в расцвете сил (10-12 лет) занимаются украшательством наиболее активно (В), к старости число украшений снова снижается (С). См. главу "Происхождение человека и половой отбор".
Беседки больших серых шалашников. Большинство красных украшений лежит справа от входа в галерею — вероятно, потому что самцу удобнее их брать оттуда во время ритуала ухаживания, чтобы показать самке. Появление культуры кловис стало итогом двухтысячелет- него развития древнего американского человечества. Возможно, центром происхождения этой культуры был юг США потому что именно здесь найдены их главные "рабочие мастерские". "Ген авантюризма" чаще встречается у кочевников, чем у оседлого населения Генетические данные показывают, что по мере того, как сапиенсы в ходе своего Великого расселения удалялись от африканской прародины, они постепенно теряли свое исходное, принесенное из Африки генетическое разнообразие. В большинстве случаев это касалось нейтральных генетических вариаций, которые терялись чисто случайно — просто в силу того, что из каждого очередного заселенного сапиенсами района в дальнейшие странствия отправлялась не вся популяция, а только ее часть. Но некоторые аллели, по-видимому, подвергались при этом действию отбора. Речь идет в первую очередь о тех генетических вариантах, которые прямо или опосредованно влияют на склонность людей к переселениям и о тех, "полезность" которых зависит от образа жизни их обладателей. Ген DRD4 является одним из самых полиморфных (вариабельных) генов в геноме человека. Белок, кодируемый этим геном, — дофаминовый рецептор D4 — работает в клетках мозга и определяет, наряду с другими белками, чувствительность определенных групп нейронов к "веществу удовольствия" — нейромедиатору дофамину (см. главу "Душевная механика", кн. 2). Аллели гена DRD4, имеющиеся в генофонде человечества, отличаются друг от друга не только многочисленными однонуклеотидными заменами, но и числом копий повторяющегося участка длиной в 48 пар нуклеотидов. Этот участок может быть повторен от двух до и раз. Соответствующие аллели (точнее, группы аллелей) называются 2R, 3R, 4R,... 11R. Чаще всего встречается варианте четырьмя повторами 4R), который является исходным, предковым вариантом данного гена у Homo sapiens. Немного реже встречаются аллели 2R и 7R. Известно, что рецептор с семью повторами (то есть белок, кодируемый аллелем 7R) реагирует на дофамин примерно вдвое слабее, чем варианты 2R и 4R. Аллель 7R в последние годы привлекает особое внимание исследователей. Его иногда называют "геном авантюризма", потому что, как оказалось, его носители имеют довольно четкие, статистически значимые поведенческие отличия от обладателей других вариантов гена DRD4. У носителей аллеля 7R в среднем сильнее, чем у других людей, выражено стремление к поиску новых ощущений, для них характерна повышенная импульсивность. Кроме того, среди них чаще встречаются люди с так называемым синдромом дефицита внимания и гиперактивности. Необходимо помнить, что термин "ген авантюризма" (как и все подобные термины) вовсе не означает, что обладание аллелем 7R является необходимым и достаточным условием формирования такого сложного фенотипического признака, как авантюризм. Это всего лишь общепринятый генетический жаргон. Любой сложный фенотипический признак формируется в результате совместной работы множества генов и факторов среды (таких, например, как воспитание). Просто гену DRD4 так "повезло", что именно у него имеются аллельные варианты, обладатели которых статистически отличаются друг от друга по степени выраженности данного признака (см. главу "Генетика души", кн. 2). Частота встречаемости аллеля 7R в разных человеческих популяциях варьирует от о до 78 %. В географическом распределении этого аллеля обнаружены крайне интересные закономерности. Чаще всего он встречается у американских индейцев, реже всего — у жителей Восточной Азии. Из европейских народов наибольшая доля носителей 7R характерна для ирландцев — народа, славящегося своей импульсивностью. Предполагают, что в Восточной Азии, где издавна существовали мощные централизованные государства, поведенческие признаки, характерные для носителей аллеля 7R, не давали преимуществ и были скорее вредны. В обществах индейцев, особенно кочевых охотников-собирателей, наоборот, гиперактивность, импульсивность и активное поисковое поведение, возможно, могли давать адаптивное преимущество. Кроме того, прослеживается положительная корреляция между частотой встречаемости 7R и расстоянием от главных азиатских центров расселения до нынешнего ареала популяции. Возможно, это означает, что повышенное число "авантюристов" в племени способствовало далеким миграциям. Или в ходе далеких миграций носители 7R получали адаптивное преимущество и размножались эффективнее, чем люди с более уравновешенным нравом. Или каждый раз, когда надо было решать, поселиться здесь или идти дальше, носители 7R отправлялись в путь, а прочие оставались. Для проверки этих увлекательных гипотез необходимы большие массивы данных по генетике различных человеческих популяций. Для строгих и окончательных выводов данных до недавних пор не хватало. Так, изменчивость гена DRD4 до сих пор была подробно изучена только в пяти популяциях североамериканских и 15 популяциях южноамериканских индейцев. При этом оставались "непокрытыми" некоторые ключевые территории, такие как Центральная и Южная Бразилия. Недавно бразильские генетики опубликовали новые данные по частотам аллелей DRD4 у трех коренных народов, проживающих в Центральной и Южной Бразилии: каинганг, гуарани-ньяндева и гуарани-кайова. Тем самым количество исследованных южноамериканских популяций было доведено до 18, а географическое "покрытие" континента стало достаточно равномерным, чтобы проверить одну из ключевых гипотез о связи между частотой встречаемости аллеля 7R и образом жизни народа (Tovo-Rodrigues et al., 2010). Давно предполагалось, что склонность к рискованному "поиску новизны" (что характерно для носителей 7R) дает преимущество в условиях нехватки ресурсов или в сильно переменчивой среде. Именно в таких условиях существует большинство охотников-собирателей. Те же самые особенности поведения могут оказаться вредными для оседлых народов, практикующих интенсивное сельское хозяйство: их источники пищевых ресурсов куда более надежны и стабильны, и поэтому "здоровый консерватизм" им в общем случае более выгоден, чем импульсивные метания в поисках чего-то нового. Проанализировав все собранные к настоящему времени данные по южноамериканским индейцам, авторы получили результат, подтверждающий эту гипотезу. Оказалось, что частота встречаемости 7R заметно выше в популяциях, которые вплоть до недавнего времени вели образ жизни охотников-собирателей (десять популяций из 18), по сравнению с сельскохозяйственными племенами (восемь популяций). Данных по североамериканским индейцам пока недостаточно для столь же достоверных выводов. Однако известно, что в Северной Америке частота встречаемости 7R несколько ниже, чем в Южной, и в целом ближе к тому, что наблюдается у оседлых (а не кочевых) южноамериканских народов. По- видимому, это связано с тем, что заселение Южной Америки было сопряжено с "бутылочным горлышком" численности и четкой дифференцировкой по темпераменту: лишь очень немногие из обитателей Северной Америки отправились заселять дикие просторы южного континента, и среди этих переселенцев, очевидно, преобладали носители "гена авантюризма". В дальнейшем, когда некоторые южноамериканские народы перешли к оседлости и производящему хозяйству, вектор отбора изменился и преимущество получили носители "неавантюрных" вариантов гена DRD4; в результате частота встречаемости 7R у таких народов снизилась. Для окончательной проверки гипотезы об адаптивном значении аллельных вариантов DRD4 и об их избирательном распространении под действием отбора необходимо выяснить, действительно ли при кочевом охотничье-собирательском укладе носители 7R лучше живут (и оставляют в среднем больше потомков), чем носители других аллелей, тогда как у оседлых земледельцев все должно быть наоборот. Подобное исследование было проведено пока только одно, и не в Америке, а в Африке. Его объектом была народность ариаал, проживающая в северной Кении. Полученные результаты подтвердили теоретические ожидания: оказалось, что в группах, ведущих традиционный кочевой образ жизни, носители 7R питаются в среднем лучше носителей других аллелей, тогда как в группах, перешедших к оседлости, ситуация сменилась на обратную (Eisenberg et al., 2008). Дополнительные подтверждения того, что аллель 7R действительно влиял на приспособленность людей и распространялся под действием отбора (а не за счет случайного дрейфа), были получены биоинформационными методами, то есть путем компьютерного анализа нуклеотидных последовательностей. Надежными признаками того, что тот или иной участок ДНК поддерживался отбором и быстро распространялся, являются повышенная доля значимых нуклеотидных замен по отношению к синонимичным заменам, а также низкий генетический полиморфизм в окрестностях рассматриваемого локуса (подробнее об этом говорилось в главе "Мы и наши гены"). Оба эти признака характерны для аллеля 7R. Биоинформационный анализ позволил заключить, что этот аллель появился в результате мутации 60 тыс. лет назад или раньше. Его первичное распространение в человеческих популяциях, вероятно, было связано с такой грандиозной "авантюрой", как выход сапиенсов из Африки и их великое расселение. Судьба "гена авантюризма" — отличный пример того, как эволюция поведенческих признаков под действием естественного отбора продолжалась в человеческих популяциях еще в недавнем прошлом, а возможно, продолжается и в наши дни. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||