|
||||
|
|
ГЛАВА 6. НАПРАВЛЕННОСТЬ ЭВОЛЮЦИИСлучайность и закономерностьОдно из центральных утверждений синтетической теории эволюции (СТЭ) состоит в том, что все эволюционные изменения основаны на отборе случайных, ненаправленных мутаций. Однако эволюция совсем не похожа на случайный, хаотический процесс. В ней явно есть направленность. Например, очевидно, что господствующие на Земле формы жизни постепенно становятся все более сложными. Как же так? Может ли направленный и как будто вполне осмысленный процесс идти на основе случайных, хаотических наследственных изменений — мутаций? В более общей форме этот вопрос можно сформулировать так: каково соотношение случайности и закономерности в эволюции? Этот вопрос был и остается одним из центральных в теоретической биологии и одновременно одним из самых спорных. Отчасти мы уже ответили на него в главе 4, где говорилось, в частности, о том, что естественный отбор — то есть избирательное запоминание системой возникающих изменений — неизбежно придает эволюции упорядоченный, направленный характер. Особенно ярко проявляется закономерный характер эволюции в явлении параллелизма. Так называют те случаи, когда у разных организмов независимо возникают похожие признаки. Мы уже вскользь упоминали об этом явлении, когда говорили о приспособлении бактерий к антибиотикам (глава 4). Пришло время поговорить о параллелизмах подробнее. Некоторые примеры общеизвестны. Так, представители разных классов позвоночных животных, перешедшие к активному плаванию в толще воды, независимо друг от друга приобрели обтекаемую рыбообразную форму тела и плавники (рыбы, ихтиозавры, дельфины). Еще более интересны примеры возникновения почти идентичных наборов "жизненных форм" на разобщенных континентах. Например, Южная Америка долгое время была изолирована от других материков, но еще до отделения туда проникли примитивные плацентарные млекопитающие кондиляртры — "древние копытные". Эволюция кондиляртр протекала далее независимо в Южной и Северной Америке, причем последняя время от времени соединялась с Евразией сухопутным мостом — Берингией. В результате на "Большой Земле" (в Северной Америке и Евразии) кондиляртры дали начало отрядам парно- и непарнокопытных, куда относятся всем известные лошади, носороги, антилопы, олени, гиппопотамы и т. д. В Южной Америке расцвели другие отряды копытных, произошедшие от тех же кондиляртр — нотоунгуляты (Нотоунгуляты, по-видимому, появились в Азии, а оттуда проникли в Северную и Южную Америку еще от ее отделения. Но на "Большой Земле" они быстро вымерли, вытесненные более совершенными группами, а в Южной Америке сохранились, размножились и дали много разнообразных, удивительных форм), литоптерны, астрапотерии и пиротерии. Среди этих "южных копытных" появились формы, поразительно схожие с настоящими лошадьми, носорогами, верблюдами, зайцами и даже слонами. В Южной Америке процветали также и сумчатые, быстро вымершие на "Большой Земле". Среди южноамериканских сумчатых тоже появились жизненные формы, удивительно похожие на некоторых североамериканских и азиатских зверей. Один из ярких примеров — сумчатый саблезубый тигр тилакосмилус, аналог настоящих саблезубых тигров, которые относились к семейству кошачьих и охотились на крупных толстокожих копытных на просторах Азии, Африки и Северной Америки. Тилакосмилус, хоть и был сумчатым, вероятно, занимал ту же экологическую нишу и охотился на крупных толстокожих "южных копытных" (Увлекательная история эволюции южноамериканских животных описана в знаменитой книге Дж. Симпсона Великолепная изоляция. http://rogov.zwz.ru/Macroevolution/simpson1983.djvu.). Впрочем, не стоит преувеличивать это сходство. В Южной Америке появились и такие звери, аналогов которым среди обитателей "Большой земли" найти не удается. Это броненосцы, муравьеды, ленивцы, к которым относятся и удивительные гигантские наземные ленивцы мегатерии. Все эти звери не похожи ни на кого, только на самих себя. В целом южноамериканские звери были менее совершенными, чем их аналоги с "Большой Земли". Например, сумчатые хищники вроде тилакосмилуса все-таки не могли сравниться в ловкости и сообразительности с "настоящими", несумчатыми, львами и тиграми. Поэтому с южноамериканскими хищными млекопитающими успешно конкурировали гигантские хищные нелетающие птицы фороракосы. Кроме того, там появились самые крупные летающие птицы всех времен. Некоторые из них, такие как аргентавис, достигали в размахе крыльев семи с половиной метров при весе до 80 кг и, возможно, могли проглотить целиком зверя размером с зайца. Совсем недавно в Патагонии был найден самый большой в мире птичий череп. Его длина — 71,6 см.
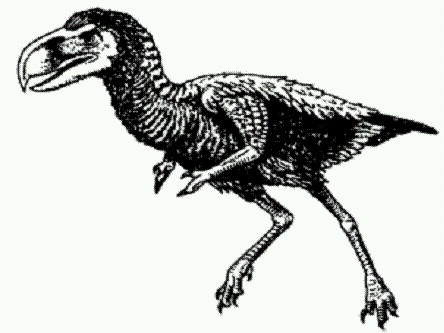
Самый большой птичий череп всех времен. Фороракосы — крупные нелетающие хищные птицы, появившиеся в Южной Америке около 62 млн лет назад, то есть почти сразу после вымирания динозавров. Многие палеонтологи полагают, что между этими двумя событиями существует тесная связь. Исчезновение динозавров освободило нишу двуногого бегающего хищника, которая активно эксплуатировалась разнообразными хищными динозаврами в течение всей истории этой группы рептилий. После вымирания динозавров эта ниша была немедленно освоена их ближайшими родственниками — птицами. Семейство фороракосов, последние представители которого вымерли около 2 млн лет назад, включает самых крупных птиц всех времен. Их рост мог достигать трех метров и более. Однако многие детали строения крупных фороракосов остаются неизвестными из-за фрагментарности имеющихся находок.
Titanis walleri — родственник фороракоса, живший 2 млн лет назад в Техасе и Флориде. На передних конечностях у него имелась пара пальцев, образующих подобие клешни. В 2006 году палеонтологи нашли в Аргентине неплохо сохранившийся череп фороракоса длиной 716 мм. Это самый большой птичий череп из всех известных науке. Возраст находки — 10-15 млн лет (средний миоцен). Вместе с черепом найдены кости ног, которые оказались неожиданно длинными и тонкими. Это противоречит прежним представлениям о том, что крупнейшие представители фороракосов были тяжеловесны и медлительны. По всей видимости, эти чудовищные птицы на самом деле были очень быстрыми бегунами. (Источник: Luis М. Chiappe, Sara Bertelli. Skull morphology of giant terror birds // Nature. 2006. V. 443.) Несмотря на удивительное разнообразие и кажущуюся высокую приспособленность южноамериканских зверей, многие из них не выдержали конкуренции с североамериканскими переселенцами: после воссоединения Южной и Северной Америк, а это произошло в плиоцене, многие северные виды отправились осваивать южные территории. Большинство южноамериканских хищников и копытных были вытеснены пришельцами и вымерли. В этом проявилось одно из "правил" эволюции, которому пока нет однозначного объяснения. На больших территориях быстрее идут эволюционный прогресс и рост конкурентоспособности, тогда как на малых территориях, в том числе на изолированных островах, быстрее происходит появление всевозможных странных, необычных, причудливых форм. Это обстоятельство биологи подметили уже давно, но лишь недавно сумели его подтвердить строгими цифрами. Эволюция на островах идет быстрее. Еще со времен Дарвина, исследовавшего галапагосских вьюрков, удаленные острова считаются своеобразными "эволюционными лабораториями", в которых виды претерпевают быстрые и порой весьма изощренные эволюционные изменения. В частности, известно, что крупные животные, попав в изолированные и обедненные островные экосистемы, часто мельчают, а мелкие, наоборот, достигают гигантских размеров, причем в ряде случаев на это требуется совсем немного времени. Однако до сих пор представления об ускоренной эволюции на островах не имели строгого количественного обоснования, а отдельные факты и вовсе им противоречили: например, ископаемому сицилийскому слону Elephas falconeri, рост которого был менее метра, а масса (100 кг) составляла лишь 1% от массы его континентального предка, потребовалось очень много времени (около 300 тысяч лет), чтобы так измельчать. Чтобы точно измерить темпы эволюционных изменений на островах и сравнить их с аналогичными данными по материковым популяциям, необходимо было собрать весьма значительный объем палеонтологических данных, причем весь материал должен был быть точно датирован. К настоящему времени палеонтология уже накопила достаточно данных для такого анализа, что и позволило Вирджинии Миллиен из Университета Макгилла (Канада) получить строгие оценки темпов эволюции на островах и на "Большой Земле".
Гигантский варан, обитающий на островах Комодо, Ринджа, Падар и Флорес (Индонезия) — один из общеизвестных случаев «островного гигантизма». Миллиен обработала данные по 86 островным и 84 континентальным популяциям млекопитающих, относящихся к 88 видам и 14 отрядам. Для каждой популяции была подсчитана скорость эволюционных изменений одного или нескольких размерных признаков (от 1 до 57 признаков для каждой популяции, в среднем 5,06 признака на популяцию) в течение определенного интервала времени (от 20 до 12 000 000 лет, средняя величина интервала — 462 тысячи лет). Использовались только линейные признаки (длина черепа, отдельных костей, зубов и т. д.). Выяснилось, что морфологическая эволюция на островах действительно идет быстрее, однако этот эффект заметен лишь на временных интервалах менее 45 тысяч лет. При увеличении временного интервала выше этого порога различия между островными и континентальными популяциями становятся статистически незначимыми. Выявленные различия нельзя объяснить разным таксономическим составом островной и континентальной биоты (то есть тем, что на островах некоторые группы млекопитающих встречаются чаще, чем на континенте, и наоборот). Оказалось, что темпы эволюции не являются "филогенетически консервативными". Например, виды, эволюционировавшие на континенте медленнее других, вовсе не обязательно будут продолжать вести себя сходным образом и на изолированном острове. По-видимому, млекопитающие, попавшие на удаленные острова, очень быстро приспосабливаются к новым условиям, что порой сопровождается разительными изменениями размеров и пропорций тела. Из-за высокой скорости этих изменений в большинстве случаев палеонтологам не удается обнаружить переходных форм между островными эндемиками и их континентальными предками. После относительно короткого периода быстрой адаптации темп эволюции островных животных, как правило, замедляется (поэтому не удалось выявить различий в темпах эволюции на временных интервалах свыше 45 тысяч лет). Фрагментация естественных местообитаний (в том числе и в результате деятельности человека) может ставить животных в условия, аналогичные попаданию на изолированные острова. В этом случае можно ожидать резкого ускорения эволюционных изменений по "островному" сценарию. Реальность таких явлений подтверждается фактами. Например, было показано резкое ускорение эволюционных изменений размеров тела за последние 200 лет у 25 видов млекопитающих в Дании. И это явление связывают с фрагментацией местообитаний. Полученные результаты показывают, что многие виды континентальных млекопитающих потенциально способны к очень быстрым эволюционным преобразованиям. Темп эволюции может возрасти в три и более раза всего за пару десятилетий. Получается, что в условиях растущей антропогенной нагрузки на природные экосистемы многие привычные нам животные могут начать изменяться самым неожиданным и быстрым образом. (Источник: Milleien V. 2006. Morphological Evolution Is Accelerated among Island Mammals.// PLoS Biol 4 (10): e321.) Но вернемся к теме параллелизмов. В последние десятилетия существенно изменились представления о том, каким образом происходили в ходе эволюции крупнейшие прогрессивные изменения — ароморфозы (По определению крупнейшего отечественного эволюциониста И. И. Шмальгаузена, ароморфоз — это "расширение жизненных условий, связанное с усложнением организации и повышением жизнедеятельности"). Оказалось, что практически во всех хорошо изученных случаях переход на более высокий уровень организации происходит не в какой-то одной эволюционной линии, а в нескольких, развивающихся параллельно. При этом отдельные признаки, из которых складывается ароморфоз, иногда появляются в разных линиях почти одновременно, а иногда — в разное время и даже в разном порядке. Прогрессивные признаки постепенно накапливаются, пока наконец в одной или немногих линиях они не соберутся все вместе, и тогда палеонтологи "объявляют" о рождении новой группы. В этом ярко проявляется закономерный характер эволюции. В какой-то момент словно начинает носиться в воздухе новая "идея" — например, идея млекопитающих. И многие разные группы не сговариваясь начинают развиваться в одном и том же направлении, хотя и немножко разными путями. Чтобы подчеркнуть это обстоятельство, многие биологи в последние годы стали называть происхождение млекопитающих "процессом маммализации зверообразных рептилий", происхождение птиц — "орнитизацией", амфибий — "тетраподизацией", членистоногих — "артроподизацией", покрытосеменных растений — "ангиоспермизацией" и т. д. Происхождение членистоногих — "артроподизация" (вендский и кембрийский периоды)В "домолекулярную эру" в распоряжении ученых было три научных дисциплины, при помощи которых можно было реконструировать эволюционную историю организмов: 1. сравнительная анатомия, 2. сравнительная эмбриология и 3. палеонтология. В последние десятилетия добавился еще один чрезвычайно мощный метод, основанный на сравнении нуклеотидных последовательностей ДНК и аминокислотных последовательностей белков. Молекулярный метод сильно потеснил первые две дисциплины (но не третью), потому что он позволяет работать одновременно с огромным количеством признаков. По сути дела, каждый нуклеотид в молекуле ДНК — это отдельный признак, который можно использовать в эволюционных исследованиях наряду с такими классическими признаками, как строение черепа или ранние стадии развития эмбриона. Еще в конце XIX — начале XX века анализ эмбриологических и анатомических данных привел к появлению двух основных гипотез происхождения членистоногих. К этому типу животных относятся ракообразные, насекомые, многоножки, паукообразные и трилобиты. Согласно первой из них, предки членистоногих были похожи на типичную личинку ракообразных — науплиус, тело которого состоит из трех сегментов. Вторая гипотеза, ставшая в какой-то момент почти общепризнанной, предполагала, что предками членистоногих были многосегментные животные с множеством однотипных парных конечностей, близкие к современным многощетинковым кольчатым червям — полихетам. Происхождение членистоногих от кольчатых червей казалось большинству ученых совершенно очевидным. Слишком уж бросается в глаза сходство многоножки или гусеницы с кольчатым червем, и даже состоящие из двух "веточек" ножки примитивных членистоногих (ракообразных, трилобитов) удивительно похожи на двуветвистые конечности полихет. В замечательном учебнике В. А. Догеля "Зоология беспозвоночных", по которому училось несколько поколений отечественных биологов (включая автора этих строк) и продолжают учиться до сих пор, написано буквально следующее: "Происхождение типа Arthropoda в общих чертах ясно. Предками их были примитивные полимерные кольчатые черви из класса многощетинковых (Polychaeta). Сходство плана строения членистоногих и кольчецов настолько велико, что некоторые современные зоологи предпочитают объединять их, как это делали свыше 100 лет назад, в один тип членистых — Articulata (В. А. Догель. Зоология беспозвоночных. Седьмое издание. М.: Высшая школа, 1981. с. 430. http://rogov.zwz.ru/Macroevolution/dogel.djvu.). Такая степень уверенности в вопросе о том, кто от кого произошел, встречается у Догеля ох как нечасто. И надо же такому случиться — из множества гипотез, основанных на данных анатомии и эмбриологии, именно эта, по всей видимости, оказалась неверной. Ни палеонтологические, ни молекулярные данные не дали подтверждения этой стройной и логичной теории. Вместо ожидаемых переходных форм между полихетами и членистоногими были обнаружены весьма причудливые создания со странными сочетаниями признаков. Поначалу биологов "обнадежила" сприггина — мягкотелое животное вендского периода (630-542 млн лет назад; см. главу "Животные"), которое вроде бы имело ожидаемое сочетание признаков: цельную "голову" в форме полумесяца, напоминающую головной щит трилобита, и длинное тело, состоящее из одинаковых сегментов с длинными придатками, напоминающее тело многощетинковых червей. Но если внимательно присмотреться к отпечаткам сприггины, можно заметить, что конечности у нее располагаются не попарно друг напротив друга, как у всех без исключения кольчатых червей и членистоногих, а в шахматном порядке (как если бы левую половину ее тела сдвинули относительно правой на пол сегмента вперед или назад). Такая "билатеральная симметрия со сдвигом" специалистам по сравнительной анатомии казалась совершенно немыслимой. Непонятно, как росло тело сприггины, как формировались сегменты (и вообще, можно ли их назвать сегментами?), как были устроены ее кровеносная и нервная системы, структура которых определяется у полихет механизмом формирования сегментов. Еще лучше эта "невероятная" сегментация видна у других вендских форм, похожих на примитивных членистоногих, — например, у вендии.
Сприггина (вендский период) могла бы считаться переходной формой между кольчецами и членистоногими, если бы имела нормальную билатеральную симметрию.
Вендия — «членистоногое» с ногами в шахматном порядке. Многие другие вендские животные, напоминающие кольчатых червей или примитивных членистоногих, "сдвинуты" таким же образом. Состоят ли они в родстве с настоящими кольчецами и членистоногими или те произошли от каких-то других предков? Возможно ли эволюционное превращение "сдвинутой" симметрии в нормальную? Окончательных ответов пока нет. Вот еще одно животное со сдвинутой симметрией — дикинсония. Найден отпечаток ее пищеварительной системы, и видно, что отростки кишечника тоже располагались в шахматном порядке, в соответствии с сегментацией. Ясно, что это и не членистоногие, и не полихеты. Палеонтолог М. А. Федонкин выделил этих животных в особый вымерший тип проартикулят (Proarticulata).
Еще одно животное со "сдвинутой" метаметрией — дикинсония. Справа — ее пищеварительная система. По изображению из книги Я. Е. Малаховской и А. Ю. Иванцовой Вендские жители Земли, http://evolbiol.ru/vend.htm.) Забавно, что очень похожая сегментация имеется и у прикрепленных, стебельчатых вендских животных — петалонам, которые уже совсем не похожи ни на кольчецов, ни на членистоногих. Что ж, если вендская мягкотелая фауна дала больше новых загадок, чем ответов на старые вопросы, может быть, уникальные находки более поздних кембрийских мягкотелых животных скажут нам больше? Кембрийский период (488-542 млн лет назад) был временем становления большинства важнейших групп многоклеточных животных. Как мы помним из главы "Животные", в самом начале кембрия произошла скелетная революция — в палеонтологической летописи внезапно появляется множество разнообразных существ, обладающих твердыми скелетными элементами: различными раковинками, шипами, спикулами и т. д. В венде животные были в основном мягкотелыми, и среди них мало кого можно с уверенностью отнести к тому или иному известному типу. Но уже в первой половине кембрия разнообразие животных резко выросло, и среди них появились представители практически всех современных типов (членистоногие, моллюски, брахиоподы, хордовые, иглокожие и т. д.).
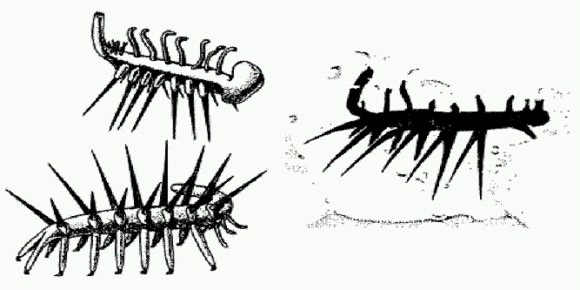
"Перистое" вендское ископаемое — чарния. Одно из самых богатых местонахождений кембрийских мягкотелых называется сланцы Берджес (Burgess Shale) и находится в Канаде. То, что было там найдено, привело ученых в полное замешательство. Чего стоят одни названия, которые ученые дали этим "возмутительным" тварям. Ведь они снова, как и вендские существа, не пожелали вписываться в красивые и стройные схемы сравнительных анатомов и эмбриологов! Одно из животных получило название "галлюцигения". Палеонтологи, видимо, долго протирали глаза, впервые увидев отпечаток этого существа. На первых найденных отпечатках было видно по три конечности на каждом сегменте: два твердых "шипа" и одно мягкое "щупальце". Правда, вендской "симметрии со сдвигом" здесь уже не было, симметрия галлюцигении — вполне правильная, билатеральная, но еще неизвестно, что хуже — ноги в шахматном порядке или по три ноги на сегменте! Сначала предполагали, что галлюцигения ходила на парных "шипах", а "щупальца" в один ряд располагались у нее на спине. Потом, к счастью, нашлись другие отпечатки, на которых удалось разглядеть, что мягких щупалец на каждом сегменте было все-таки два, а не одно. Так появилась более правильная (по крайней мере, более правдоподобная) реконструкция галлюцигении. Животное перевернули вверх тормашками и поставили на парные щупальца, а шипы стали просто защитными придатками вроде игл морского ежа. В таком виде галлюцигения напоминает, хоть и отдаленно, современных онихофор — червеобразных животных с ножками, родственных членистоногим.
Галлюцигения.
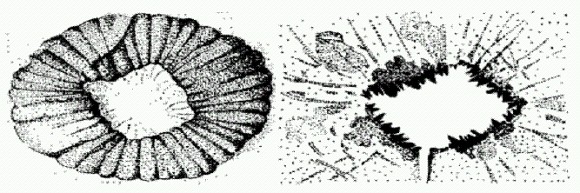
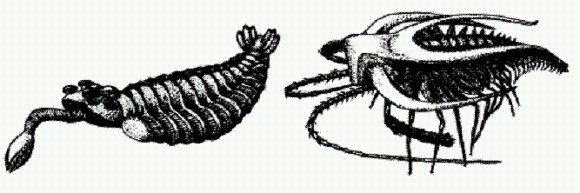
Айшеайя. Голова находится справа, видны приротовые придатки. Возможно, айшеайя питалась губками, потому что ее остатки часто находят вместе с остатками губок. В сланцах Берджес нашли и других онихофор, больше похожих на современных (Aysheaia). Но все "приключения" галлюцигении меркнут по сравнению с уникальной судьбой другого среднекембрийского монстра — аномалокариса. Отдельные части тела этого хищного чудовища (некоторые экземпляры достигали почти двух метров в длину) сначала находили по отдельности и описывали как разные организмы. Удивительная "медуза" с дыркой посередине оказалась ротовым диском аномалокариса. Внутренний край "дырки", как выяснилось, был усажен острыми зубами. Членистые образования, изначально описанные как креветки, оказались хватательными конечностями необычного хищника. Только когда были найдены первые полные отпечатки аномалокариса, ученые поняли, как было устроено это существо. Сегментированное тело с "плавательными лопастями" напоминает некоторых кольчатых червей. Однако передние хватательные конечности (бывшие "креветки") и крупные глаза на стебельках — в точности как у членистоногих. Мягкотелая фауна сланцев Берджес и других подобных местонахождений в Китае, США, Гренландии и Сибири открыла перед учеными целую галерею удивительных существ с разнообразными причудливыми сочетаниями признаков членистоногих и червей (причем не кольчатых, а других — так называемых головохоботных). Все это очень мало похоже на постепенный, плавный переход от одного типа к другому. Вот, например, опабиния (вверху слева) — животное, явно близкое к аномалокарису, но лишенное членистых приротовых конечностей. Вместо них у опабинии имеются складной хоботок и пять глаз.
Аномалокарис — медуза с отверстием? Справа видны зубы, окружавшие отверстие.
"Креветки*.
Другие загадочные существа из Берджес-Шелл: опабиния (слева) имарелла (справа). По-видимому, отдельные признаки членистоногих формировались независимо в разных группах червеобразных организмов. Эти признаки перекомбинировались и смешивались, образуя самые неожиданные комбинации. Возможно, не обошлось здесь и без горизонтального межвидового обмена генами (такой перенос могли осуществлять вирусы). Как мы помним из прошлой главы, большую роль в крупных эволюционных перестройках у животных могут играть мутации ключевых генов — регуляторов развития, таких как Нох-гены. В результате таких мутаций свойства одних сегментов могут проявиться у других. Теоретически в результате подобной мутации от организмов, похожих на аномалокариса (с единственной парой членистых конечностей, развившихся специально для схватывания добычи), могли буквально в одночасье произойти формы, имеющие такие конечности на всех сегментах туловища. Что же касается кольчатых червей, то они, судя по современным молекулярным данным, вовсе не являются предками членистоногих. Вместе с головохоботными и круглыми червями членистоногих теперь объединяют в надтип "линяющих" Ecdysozoa, а кольчатых червей вместе с моллюсками и брахиоподами поместили в другой надтип — Lophotrocbozoa. Новые палеонтологические находки в сланцах Берджес и других местонахождениях помогают выяснять детали ранней эволюции этих групп. Почти каждая публикация такого рода служит новым подтверждением общего правила мозаичного распределения признаков и многочисленных параллелизмов на ранних этапах развития больших групп организмов. Происхождение неправильных морских ежей (юрский период)Эволюционные переходы от одной крупной группы организмов к другой, сопровождающиеся многочисленными параллелизмами и мозаичным распределением признаков, характерны не только для таких древних времен, как вендский и кембрийский периоды. В этом разделе мы перенесемся на несколько сотен миллионов лет вперед — в юрский период мезозойской эры — и рассмотрим один из важнейших переломных моментов в эволюции класса морских ежей (Echinoidea). Морские ежи делятся на "правильных" — радиально-симметричных и "неправильных" — двусторонне-симметричных. Правильные ежи всеядные, живут на поверхности грунта, а неправильные зарываются в грунт и питаются детритом, то есть органическими остатками, которыми изобилует морское дно. Доказано, что неправильные ежи произошли от правильных в первой половине юрского периода. Это, несомненно, был ароморфоз, хотя и не такой значительный, как появление млекопитающих или членистоногих. Все признаки ароморфоза налицо: строение животных усложнилось, разнообразие жизненных условий расширилось, так как был освоен принципиально новый тип субстрата — рыхлые грунты, а также обретен новый тип питания; кроме того, это эволюционное преобразование открыло морским ежам путь к увеличению видового разнообразия, численности и биомассы, то есть к тому, что называют "биологическим прогрессом". На пути от правильных ежей к неправильным ясно видны направленность движения и возможность достижения конечной цели несколькими дорогами. У правильных морских ежей анальное отверстие (перипрокт) расположено на вершине панциря и окружено десятью пластинками апикальной системы, от которых радиально расходятся двойные ряды пластинок — амбулакры и интерамбулакры. Рот находится в середине нижней стороны тела.
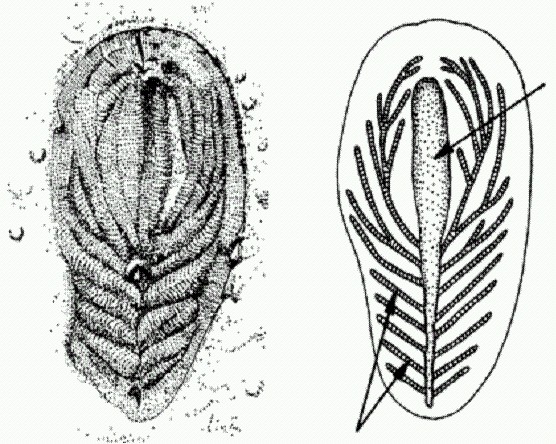
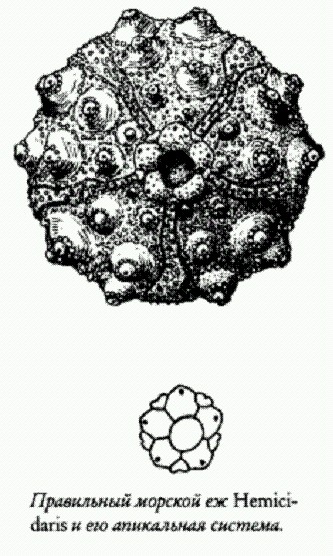
Правильный морской еж Hemicidaris и его апикальная система. В юрском периоде в нескольких группах правильных ежей независимо возникла тенденция к смещению перепрокта назад. Это было связано с переходом к питанию грунтом. При этом резко увеличился объем пропускаемого через кишечник материала, а значит, и количество отбросов. Морским ежам — грунтоедам крайне невыгодно иметь перипрокт на макушке панциря, потому что пропущенные через кишечник частицы грунта будут падать на них сверху и с большой вероятностью снова заглатываться. Для грунтоедов, и без того сидящих на низкокалорийной диете, это совершенно неприемлемо. Смещение перипрокта назад требовало изменения конфигурации окружающих перипрокт пластинок. Эти перестройки неизбежно вели к преобразованию всей архитектуры панциря, поскольку апикальные пластинки в ходе онтогенеза "отпочковывают" от себя все остальные пластинки панциря.
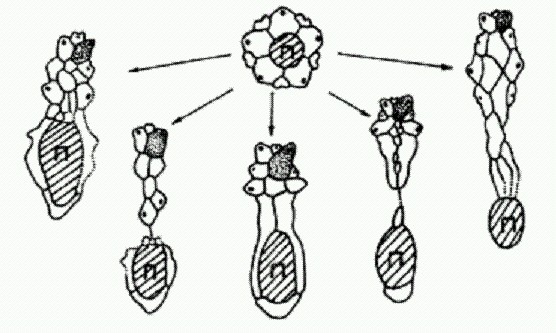
Смещение перипрокта назад — "преадаптация" к превращению морских ежей из "правильных" в "неправильных". Устоявшаяся древняя связь между перепроктом и апикальной системой не могла быть сразу отменена. Природа изобретала различные конструктивные решения этой задачи. Потребность сместить перипрокт подальше назад приводила к появлению разнообразных, в том числе весьма необычных, форм со всевозможными дополнительными пластинками; иногда апикальная система буквально разрывалась пополам.
Преобразования апикальной системы в ходе смещения перипрокта у разных групп морских ежей.
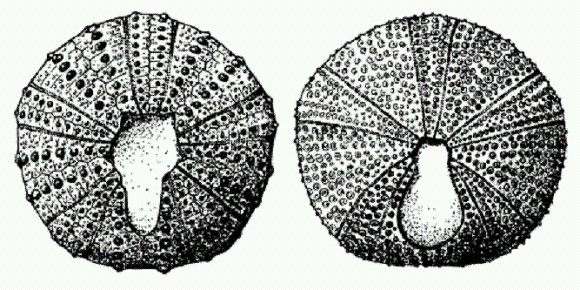
Типичный неправильный морской еж. Задняя половинка апикальной системы в итоге все-таки отделилась от перипрокта и воссоединилась с передней половиной. Апикальная система снова "собралась" вместе и вернулась в свое исходное положение на макушке панциря, а перипрокт оказался сдвинут на задний конец. В итоге окончательное отделение перипрокта от апикальной системы произошло параллельно как минимум в пяти разных эволюционных линиях (Н. Соловьев, А. В. Марков. Ранние этапы эволюции неправильных морских ежей. 2004. http://evolbiol.ru/sborniko4/o4markov.htm.). Мозаичное распределение примитивных и прогрессивных признаков, единая общая направленность изменений и многочисленные параллелизмы в ходе превращения правильных морских ежей в неправильных проявились не только в эволюции апикальной системы, но и в развитии других частей тела. Примерно таким же образом развивались у неправильных морских ежей специализированные органы дыхания, сбора и захвата пищи, передвижения в толще грунта. Происхождение птиц: "орнитизация" (конец юрского — меловой период)Мало кто из древних обитателей Земли может сравниться по своей популярности со знаменитым археоптериксом, восемь скелетов которого найдено в Германии в отложениях позднеюрского возраста. Это существо сочетает в себе признаки хищных динозавров-теропод и птиц. Пока археоптерикс был единственной известной "полуптицей-полурептилией", ситуация казалась простой. От динозавров произошел археоптерикс, от археоптерикса — настоящие птицы. Но последовали новые находки, которые, как водится, вместо того чтобы еще более прояснить ситуацию, сильно ее запутали. Во-первых, выяснилось, что археоптерикс был совсем не одинок. В отложениях мелового периода обнаружилось множество близких к археоптериксу существ, обладающих смесью птичьих и динозавровых признаков. Этих странных птиц назвали "энанциорнисами", т. е. "противоптицами", чтобы подчеркнуть, что они, возможно, и не имеют прямого отношения к настоящим птицам. Орнитизация, или "оптичивание", шла многими параллельными путями, и какая именно из "переходных форм" дала начало современным птицам, не так-то просто понять. Археоптерикс и его родичи энанциорнисы действительно очень близки к хищным динозаврам — тероподам. Хотя у них выработались многие птичьи признаки, до сих пор у ученых остались сомнения в том, что энанциорнисы были прямыми предками современных птиц. Эксперты отмечают, что у энанциорнисов не заметно тенеденции к постепенному приобретению тех птичьих особенностей, которых им "не хватало", чтобы стать настоящими птицами.
Энанциорнис. Зато обнаружились, причем в большом количестве, разнообразные наследники хищных динозавров, близкие по признакам к энанциорнисам или к каким-то другим птицеподобным созданиям. Оказалось, что многие мелкие хищные динозавры в разное время приобретали те или иные птичьи черты. Найдено довольно много видов динозавров с настоящими перьями.
Зачем динозаврам перья? Перья не сразу стали использоваться для полета. Сначала они, по всей видимости, служили для термоизоляции и были похожи на пух современных птиц. Потом они пригодились для брачных демонстраций. В октябре 2008 года в Китае нашли остатки пушистого нелетающего динозаврика размером с голубя, у которого на хвосте было четыре длиннейших пера, примерно таких же, как у самцов современных райских птиц. Такие перья могли использоваться только для привлечения самок — больше они ни на что не годны. Позже перья могли пригодиться как средство планирования при прыжках с ветки на ветку (у древесных динозавров дромеозаврид) либо для ускорения бега — у быстро бегающих наземных форм. Недавно откопали совсем уж невероятных "четырехкрылых" динозавров — микрорапторов, которые, скорее всего, умели неплохо летать. Крупные перья, явно предназначенные для полета, были у них не только на передних конечностях, но и на задних! До недавнего времени считалось, что все "оптичившиеся" динозавры были сравнительно мелкими. Однако в 2007 году в Китае был найден настоящий птицеподобный гигант, живший в позднемеловую эпоху и весивший около полутора тонн. В Китае нашли кости гигантского птицеподобного динозавра. По мнению большинства ученых, непосредственными предками птиц были целюрозавры (это одна из групп теропод, являющихся, в свою очередь, одной из групп ящеротазовых динозавров). Многие целюрозавры в течение юрского и особенно мелового периодов приобретали различные птичьи черты, в том числе перья, причем эти события происходили независимо в разных эволюционных линиях. Эксперименты по "орнитизации" продолжались и после появления настоящих птиц — по-видимому, вплоть до самого конца эпохи динозавров, наступившего на рубеже мезозойской и кайнозойской эр (65,5 млн лет назад).
Один из многочисленных видов пернатых динозавров. До сих пор считалось, что степень "оптичивания" у целюрозавров коррелировала с размерами: наибольшее число птичьих черт было отмечено у мелких представителей группы, тогда как у крупных целюрозавров орнитизация не заходила так далеко. Предполагалось, что, увеличиваясь в размерах, целюрозавры могли терять некоторые птичьи признаки и возвращаться к более примитивному состоянию. Новая находка китайских палеонтологов показала, что даже крупнейшие представители группы могли быть очень похожи на птиц. Чудовище, названное гигантораптором, проживало на территории Китая в позднемеловую эпоху, в период между 89,3 млн и 65,5 млн лет назад. Оно имело 8 м в длину, 3,5 м в высоту при горизонтальном положении туловища и весило около полутора тонн. Исследовав структуру костей на срезах, исследователи пришли к выводу, что найденный динозавр погиб на 11-м году жизни (возраст определили по числу "годовых колец"). Он уже был взрослым, но продолжал расти. По-видимому, к старости гиганторапторы могли весить существенно больше полутора тонн. Как и другие гигантские динозавры, гигантораптор рос очень быстро, значительно быстрее, чем его ближайшие родственники — некрупные целюрозавры из группы Oviraptorosauria. В строении скелета гигантораптора выявлен ряд черт, которые сближают его с птицами и ранее у крупных динозавров не встречались. В частности, из всех гигантских теропод у гигантораптора были самые длинные и тонкие конечности — как передние, так и задние. Авторы находки полагают, что гигантораптор, возможно, был самым быстрым бегуном в своем размерном классе. Отпечатки перьев не сохранились, но исследователи полагают, что перья у гигантораптора могли быть, особенно на передних конечностях (предположение основано на наличии перьев у родственников и предполагаемых предков гигантораптора и на некоторых косвенных признаках). Есть мнение, что перья у бегающих теропод изначально развились для тепла и лишь потом были приспособлены для полета. Однако нам следует иметь в виду, что крупные перья на передних конечностях никогда не служили для теплоизоляции: скорее, они выполняли функции аэродинамического характера. (Источник: Xing Хu et al. A gigantic bird-like dinosaur from the Late Cretaceous of China // Nature. 2007. V. 447. P. 844-847.) Как заметил российский палеоорнитолог Е. Н. Курочкин, у археоптерикса на сегодняшний день не осталось ни одного "птичьего" признака, который не был бы найден у тех или иных динозавров. Раньше такими признаками считались перья, крючковидные отростки на ребрах и вилочка (сросшиеся ключицы). Зато многие из тех признаков, которые есть у настоящих (веерохвостых) птиц, но отсутствуют у динозавров, не обнаружены ни у археоптерикса, ни у его ящерохвостых родственников. Кто был предком настоящих птиц? Археоптерикс и его родня — энанциорнисы, — несомненно, произошли от динозавров. Однако, по мнению Е. Н. Курочкина, это была тупиковая ветвь, вымершая в конце мелового периода вместе с динозаврами и не оставившая потомков. Курочкин считает, что предков настоящих птиц нужно искать в более древних эпохах. Возможно, птицы произошли вовсе не от пернатых динозавров, не от археоптерикса и не от энанциорнисов, а от более древних пресмыкающихся — позднетриасовых текодонтов. Не исключено, что эта группа рептилий была общим предком и птиц, и динозавров. Лучшим из обнаруженных до сих пор кандидатов на роль птичьего предка среди текодонтов является протоавис, найденный в позднетриасовых отложениях США в 1983 г. Хотя протоавис был наземным животным и не умел летать, у него имеются важнейшие птичьи признаки, которых нет ни у археоптерикса, ни у энанциорнисов, ни у пернатых динозавров. Возможно, движение в "птичью" сторону началось еще в триасе у текодонтов. Линия, приведшая к настоящим птицам и идущая от форм, близких к протоавису, не сразу достигла эволюционного успеха. Вначале доминировала "энанциорнисовая" линия, идущая от хищных динозавров — теропод. Причем птичьи признаки приобретались параллельно в нескольких линиях динозавров, поэтому говорят о "процессе орнитизации теропод". Эта линия оказалась тупиковой. Только после ее вымирания в конце мела настоящие (веерохвостые) птицы заняли освободившееся экологическое пространство и достигли подлинного расцвета. Впрочем, большинство западных ученых не разделяют эту точку зрения и выводят птиц напрямую из энанциорнисов и им подобных промежуточных форм. Завершая разговор о происхождении птиц, расскажу об одном необычном исследовании, результаты которого были опубликованы в начале 2007 года в журнале Nature (Chris L. Organ, Andrew M. Shedlock, Andrew Meade, Mark Pagel, Scott V. Edwards. Origin of avian genome size and structure in non-avian dinosaurs // Nature. 2007. V. 446. E 180-184.). Кто бы мог подумать, что в серьезных научных журналах начнут появляться статьи об эволюции геномов динозавров — животных, в чьих ископаемых костях за давностью лет никакой ДНК попросту не осталось. Тем не менее это произошло. Изобретательность исследователей, выполнивших эту работу, вызывает чувство глубокого восхищения. Ученые воспользовались тем обстоятельством, что в ископаемых костях, если они достаточно хорошо сохранились, на срезах бывают видны маленькие полости, в которых при жизни животного помещались клетки костной ткани — остеоциты. Известно, что размер генома в некоторых группах живых существ положительно коррелирует с размером клеток. Справедливо ли это для остеоцитов позвоночных? Авторы изучили срезы костей 26 видов современных тетрапод (то есть четвероногих; к ним относятся амфибии, рептилии, птицы и млекопитающие) и обнаружили линейную зависимость между размером генома и средним объемом остеоцита. Найденная зависимость позволила ученым с приемлемой точностью оценивать размеры геномов ископаемых видов. Авторы воспользовались этой замечательной возможностью, чтобы ответить на вопрос, который давно волнует ученых: когда и почему у предков современных птиц произошло радикальное уменьшение генома? Дело в том, что птичьи геномы значительно меньше по размеру, чем у остальных тетрапод. Размер генома современных птиц — от 0,97 до 2,16 млрд пар нуклеотидов, в среднем 1,45. Для сравнения — у жабы 6,00, у крокодила 3,21, у коровы 3,7, у кошки 2,9, у мыши 3,3, у человека 3,5. Считалось, что уменьшение генома у птиц является своеобразной адаптацией к полету. Из генома была выброшена значительная часть некодирующих и повторяющихся участков, в частности многие мобильные генетические элементы. По логике вещей для птиц было важнее всего максимально облегчить свое тело и оптимизировать обмен веществ. Наличие в каждой клетке сотен миллионов "лишних" пар нуклеотидов было бы для них непозволительной роскошью: ведь все эти километры ДНК нужно еще и обслуживать — определенным образом упаковывать, ремонтировать при возникновении различных поломок, копировать перед каждым клеточным делением. А для этого клетка должна синтезировать и содержать огромное количество различных белковых молекул, не говоря уже об энергетических затратах. Но это только логика, а природа зачастую действует в обход человеческого разумения. Чтобы проверить, действительно ли уменьшение генома было связано с полетом, необходимо было выяснить размеры генома у вымерших предков птиц. Ученые исходили из теории о происхождении птиц от динозавров-теропод. Между прочим, в силу особенностей современной биологической систематики факт происхождения птиц от динозавров (а не от общих с динозаврами предков) требует рассматривать птиц как подгруппу динозавров, а для "собственно" динозавров теперь применяют громоздкий термин "non-avian dinosaurs" (нептичьи динозавры). То есть формально динозавры не вымерли: посмотрите в окно — на ветках сидит множество крылатых "динозавриков"! Ученые измерили остеоциты у 31 вида динозавров и ископаемых птиц, или, как они пишут, "птиц и нептичьих динозавров". По размеру остеоцитов они прикинули возможные размеры геномов их владельцев. Результаты получились довольно неожиданные. Оказалось, что размеры остеоцитов и, следовательно, геномов сильно различаются у двух основных групп динозавров — птицетазовых и ящеротазовых. К птицетазовым относятся растительноядные формы, такие как трицератопс и игуанодон. К ящеротазовым помимо двуногих хищников — теропод относятся также массивные диплодоки и им подобные из группы завропод. И как будто нарочно, чтобы всех запутать, птицы произошли вовсе не от птицетазовых, а от ящеротазовых динозавров. Как выяснилось, геномы птицетазовых динозавров имели средний размер около 2,5 млрд пар нуклеотидов, что вполне сопоставимо с современными рептилиями. Геномы теропод, в том числе самых древних, живших задолго до появления птиц, были значительно меньше — в среднем 1,78 млрд п. н. Из девяти исследованных видов теропод только у одного (овираптора) размер генома оказался за пределами диапазона, характерного для современных птиц. Единственный исследованный представитель завропод — апатозавр — тоже имел маленький геном. Авторы пришли к заключению, что общий предок всех динозавров имел типичный для наземных позвоночных большой геном. Это состояние сохранилось у птицетазовых динозавров, а также у рептилий, доживших до наших дней. У ящеротазовых динозавров еще на заре их истории (в триасе) произошло радикальное сокращение генома. Птицы, таким образом, унаследовали маленький геном от своих предков — динозавров-теропод, а не приобрели его позже как адаптацию к полету. И все таки связь между размером генома и полетом существует. Об этом свидетельствуют два обстоятельства. Во-первых, у нелетающих птиц, таких как страусы, геном больше, чем у летающих. По-видимому, утрата способности к полету привела к тому, что всевозможные мобильные элементы снова "расплодились" в геноме нелетающих птиц. Во-вторых, у летучих мышей геном меньше, чем у других млекопитающих. По-видимому, уменьшение генома у ящеротазовых динозавров следует рассматривать не как адаптацию к полету, а как «реадаптацию, то есть как признак, развившийся в связи с какими-то иными обстоятельствами, а впоследствии облегчивший развитие способности к полету. Параллельная эволюция в высыхающем Арале. Параллелизмы характерны отнюдь не только для крупных эволюционных событий, связанных с переходом на более высокий уровень организации. Поразительные примеры параллельной эволюции наблюдались в последние годы в гибнущем Аральском море. О них стало известно благодаря исследованиям биологов из Омска С. И. Андреевой и Н. И. Андреева. Как известно, сейчас Арала как единого водоема уже не существует: он разделился на два изолированных, быстро высыхающих пересоленных "озера" — Большой и Малый Арал. Резкое увеличение солености привело к вымиранию большей части видов животных и растений. Однако некоторые двустворчатые моллюски сумели выжить. Внезапная перемена условий привела к тому, что уцелевшие виды начали быстро эволюционировать. У них резко возросла изменчивость и появились целые "букеты" новых форм, причем различия между этими новыми формами и исходными видами порой очень велики: такая степень различий характерна для разных родов, а иногда и семейств двустворчатых моллюсков. Массовое вымирание освободило множество экологических ниш. Вымерли, не выдержав повышения солености, все моллюски-фильтраторы. Однако многие зарывающиеся грунтоеды оказались более "солеустойчивыми". Опустевшие ниши фильтраторов стали "втягивать" в себя выживших моллюсков, которые ранее вели совсем другой образ жизни. В результате зарывающиеся грунтоеды из рода Cerastoderma буквально на глазах изумленных исследователей стали вылезать на поверхность грунта и превращаться в фильтраторов (при этом в строении их раковины происходили соответствующие изменения). Самое интересное, что все эти эволюционные процессы протекали очень сходным образом в двух разобщенных водоемах — Большом и Малом Арале! К сожалению, этот уникальный эволюционный "эксперимент" уже закончился, и закончился трагически — как и любое событие, ставшее следствием неразумного вмешательства человека в природу. Соленость в Большом Арале недавно достигла непереносимого для моллюсков уровня в 6%, что привело к их полному вымиранию. Возможно, в Малом Арале они проживут еще какое-то время, но сравнивать их эволюцию уже не с чем. Видообразование на разных островах идет параллельными путямиУединенные острова и озера представляют собой уникальные эволюционные лаборатории, где небольшие популяции животных и растений могут сильно меняться за весьма малое по геологическим масштабам время. Этому способствует прежде всего обедненность таких изолированных экосистем и резко ослабленная межвидовая конкуренция. Всевозможные причудливые, узко специализированные формы, которые на материке, скорее всего, не имели бы никаких шансов на существование, на крошечном уединенном островке имеют возможность выжить. Изучение островных флор и фаун иногда позволяет почти воочию увидеть процесс эволюции и проверить на практике различные теоретические модели, разработанные биологами. Южноафриканские исследователи, изучающие эндемичных вьюрков архипелага Тристан-да-Кунья, в начале 2007 года опубликовали в журнале Science статью, в которой подтверждаются сразу две широко обсуждаемые в научной литературе гипотезы (Peter G. Ryan, Paulette Bloomer, Coleen L. Moloney, Tyron J. Grant, Wayne Delport. Ecological Speciation in South Atlantic Island Finches // Science. 2007. V. 315. R 1420-1423.). Первая из них — модель экологического видообразования в условиях симпатрии (то есть на одной и той же территории). Согласно модели, исходный предковый вид может разделиться на два в результате адаптации его представителей к разным экологическим нишам, причем для этого не нужны какие-либо физические барьеры, разделяющие исходно единую популяцию на две изолированные части. Замечу, что в рамках классической СТЭ симпатрическое видообразование традиционно считалось весьма маловероятным; думали, что в подавляющем большинстве случаев для разделения вида необходимы физические изолирующие барьеры. В последние годы, однако, получено множество убедительных доказательств реальности симпатрического видообразования. Вторая гипотеза предполагает, что, поскольку видообразование контролируется экологическими факторами, оно должно в сходных условиях происходить сходным образом. Иначе говоря, если есть два одинаковых изолированных острова и на каждый из них попадет один и тот же предковый вид, то через некоторое время на этих островах должны параллельно и независимо образоваться сходные "пучки" видов. Конечно, обе теоретические модели возникли не на пустом месте, и каждая из них подтверждается множеством фактов, однако в большинстве своем это косвенные доказательства. Кроме того, они затрагивают лишь отдельные группы животных или растений. Например, убедительные свидетельства симпатрического видообразования получены для пальм, многих пресноводных рыб, рачков-бокоплавов (гаммарид) озера Байкал и т. д., тогда как с другими группами (в том числе с птицами) дело обстоит хуже. К примеру, галапагосские вьюрки, получившие всемирную известность благодаря тому впечатлению, которое они произвели на молодого Чарльза Дарвина, пока не предоставили ученым убедительных свидетельств симпатрического характера своего видообразования (предполагают, что разные виды этих птиц могли сформироваться на разных островах). То же самое до настоящего времени предполагалось и для вьюрков архипелага Тристан-да-Кунья. Орнитологи делят их на два вида: Nesospiza acunhae (с маленьким клювом) и N. Wilkinsi (с большим клювом). Оба вида встречаются только на двух маленьких островах — Неприступном (Inaccessible) площадью 14 кв. км и Найтингейл (Nightingale) площадью 4 кв. км. Первый вид ранее обитал также и на главном острове архипелага, но там его истребили около ста лет назад завезенные людьми крысы и мыши. Острова Неприступный и Найтингейл, к счастью, необитаемы и к тому же объявлены заповедной зоной, так что уникальные местные птицы имеют шанс задержаться на этом свете. Между прочим, палеонтологи недавно установили, что практически на всех островах и атоллах Тихого океана до прихода людей обитали разнообразные эндемичные виды птиц, в том числе нелетающих. Как только на остров приходили люди — полинезийцы или меланезийцы, — эти виды немедленно исчезали. Так что истории с дронтом и новозеландским моа — не исключение, а правило.
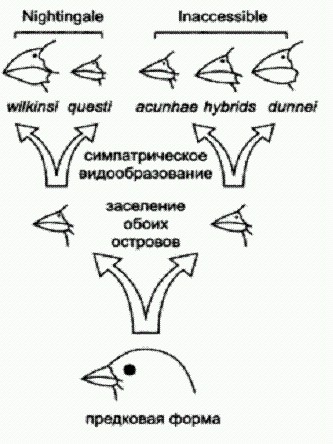
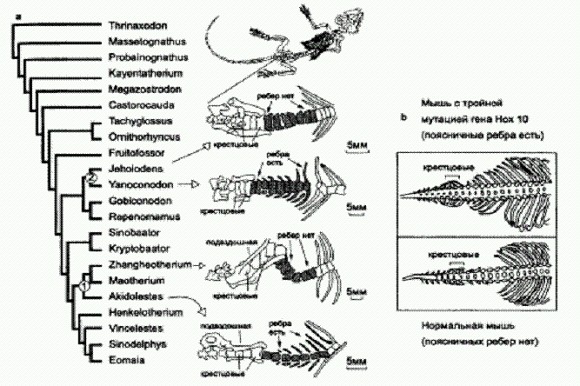
Схема эволюции вьюрков рода Nesospiza. Представители одного и того же вида, обитающие на разных островах, немного отличаются друг от друга, поэтому их считают разными подвидами. На Неприступном выделяют подвиды N. acunhae acunhae и N. wilkinsi dunnei, на Найтингейле — N. acunhae questi и N. wilkinsi wilkinsi. Предполагалось, что виды N. acunhae и N. wilkinsi сформировались на разных островах, а затем расселились, и их ареалы пересеклись. Однако генетический анализ, проведенный исследователями, показал, что история у этих птиц была иной. Сравнив несколько митохондриальных и ядерных генетических маркеров, ученые обнаружили, что два вида на каждом из островов ближе друг к другу, чем к сородичам своего же вида, но с другого острова. Например, большеклювые N. wilkinsi dunnei с Неприступного гораздо ближе к мелкоклювым N. acunhae acunhae с того же острова, чем к большеклювым N. wilkinsi wilkinsi с Найтингейла. Ученые сделали вывод, что все четыре формы, по- видимому, происходят от одного предка, когда-то залетевшего из Южной Америки с преобладающими западными ветрами (расстояние от материка — 3000 км). Эта предковая форма заселила оба острова и на каждом из них параллельно и независимо подразделилась на две разновидности — мелкоклювую и большеклювую. Направленность этих эволюционных преобразований определялась особенностями кормовой базы. На обоих островах основную пищу вьюрков составляют мелкие семена местной травы Spartina arundinacea и крупные семена дерева Phylica arborea. Размер клюва у вьюрков строго коррелирует с размером разгрызаемых семян. Большеклювые N. wilkinsi питаются в основном семенами дерева, мелкоклювые N. acunhae — семенами травы. Любопытно, что на меньшем по площади острове Найтингейл процесс симпатрического видообразования продвинулся заметно дальше, чем на более крупном острове. На Неприступном, где разнообразие природных условий выше, две формы вьюрков, по-видимому, еще способны скрещиваться друг с другом. В некоторых районах острова встречаются птицы с клювами промежуточных размеров — судя по всему, гибриды (генетический анализ не противоречит этому предположению). На Найтингейле никаких гибридов нет, да и генетически две местные разновидности вьюрков сильнее отличаются друг от друга, чем формы с Неприступного. По-видимому, это говорит о том, что разнообразие условий обитания в данном случае выступало как фактор, тормозящий эволюцию. Что такое "скорость эволюции"? Необходимо помнить, что под "скоростью эволюции" в зависимости от контекста могут подразумеваться два совершенно разных показателя. Одно дело — скорость образования разнообразных причудливых специализированных форм, совсем другое — скорость прогрессивных преобразований, связанных с выработкой новых адаптаций широкого профиля и ростом сложности организма. На маленьких изолированных клочках суши выше только первая из этих скоростей, тогда как вторая, наоборот, выше на больших материках с разнообразными условиями и сложными насыщенными экосистемами. Что заставляет эволюцию повторяться?Для начала отметим первое, что бросается в глаза. Сходство параллельно развивающихся групп обычно тем больше, чем ближе их родство. Очень далекие друг от друга группы могут выработать лишь самое поверхностное сходство (например, рыбы и дельфины). Наоборот, в Большом и Малом Арале или на крошечных островках архипелага Тристан-да-Кунья сходство вновь образующихся форм было очень велико, поскольку они происходили от одних и тех же исходных видов. По-видимому, само строение организма во многом предопределяет возможные пути его эволюции. Для каждого организма существует весьма ограниченное число "разрешенных" путей эволюционных изменений — и любые попытки уклониться от этих путей безжалостно отсекаются отбором. У близкородственных видов, индивидуальное развитие которых идет по сходной "программе", набор этих разрешенных путей тоже сходен, что и приводит к параллелизмам. Во-вторых, большую роль играет сходство структуры основных типов сообществ, или экосистем. Набор экологических "ролей" (ниш) в разных сообществах в целом сходен. Например, если это степь, прерия или саванна, то там должна быть, во-первых, трава, во-вторых — крупные травоядные звери (копытные или их аналоги), в третьих — хищники, охотящиеся на этих копытных, в четвертых — жуки-навозники и другие беспозвоночные, перерабатывающие навоз травоядных, и т. д. Все эти ниши обязательно должны быть кем-то заполнены, иначе весь комплекс развалится. Попробуйте лишить экосистему одного из блоков, например жуков-навозников, — немедленно начнется экологический кризис. Нечто подобное наблюдалось в Австралии, где завезенные европейскими фермерами овцы стали быстро приводить пастбища в негодность из-за отсутствия в местной фауне эффективных переработчиков навоза. Пришлось срочно завозить жуков-навозников с "большой земли". У фермеров не было времени ждать, пока эволюция справится с проблемой в своей обычной неторопливой манере. А подождали бы пару сотен тысячелетий — и почти наверняка на основе местных австралийских жуков развились бы "свои" эффективные навозники. И они, скорее всего, по многим признакам были бы похожи на своих коллег с других континентов. Обитание в сходных условиях и нацеленность на сходный образ жизни приводят к тому, что организмы приобретают сходные черты строения. Кроме того, в последнее время появляется все больше свидетельств, что "случайные и ненаправленные" наследственные мутации, которые, согласно господствовавшим до недавнего времени представлениям, являются единственным источником материала для отбора, в действительности часто оказываются не такими уж случайными. Об этом мы поговорим подробно в главе "Управляемые мутации". Конечно, природе постоянно приходится действовать "методом случайного тыка", но она всячески старается этот метод оптимизировать. Наконец, многие параллелизмы могут иметь довольно простые генетические объяснения. Индивидуальное развитие животных и растений контролируется сравнительно небольшим числом ключевых генов-регуляторов. Если, например, в двух эволюционных линиях независимо "сломается" какой-то регуляторный ген, в этом едва ли можно усмотреть что-то удивительное или невероятное. Однако внешнее проявление этой ошибки может показаться в высшей степени удивительным: ведь мы увидим весьма сложные и, главное, одинаковые изменения в двух разных группах организмов. Один из ярких примеров такого рода дают поясничные ребра, которые на ранних этапах эволюции млекопитающих многократно исчезали и снова появлялись в разных эволюционных линиях. Недавно китайские палеонтологи нашли очередного доисторического зверька, строение которого проливает новый свет на это явление. Данная находка, помимо прочего, представляет собой прекрасный пример классической "переходной формы". Надо сказать, что находки древних мезозойских млекопитающих в последнее время необычайно участились. Еще пару десятилетий назад считалось, что мезозойские млекопитающие были малочисленной, второстепенной группой мелких крысоподобных зверьков, влачивших жалкое существование в тени господствовавших в то время динозавров. Благодаря находкам последних лет, значительная часть которых сделана китайскими палеонтологами, стало известно, что древние звери были весьма многочисленны, разнообразны и могли достигать размеров лисы или барсука. Новая палеонтологическая находка проливает свет на раннюю эволюцию млекопитающих. Китайские палеонтологи в очередной раз удивили научный мир необычной находкой. На этот раз они откопали очень хорошо сохранившийся скелет неизвестного ранее примитивного млекопитающего из группы триконодонтов, жившего около 125 млн лет назад (в раннемеловую эпоху). Новооткрытый мезозойский зверек Yanoconodon allini был как раз таким, каким "положено" быть мезозойскому зверю — маленьким и невзрачным. Интересен он прежде всего тем, что у него очень хорошо сохранились косточки среднего уха (молоточек и наковальня), причем сохранились они не по отдельности, а в своем естественном положении, сочлененные с другими костями. Строение этих косточек является одним из главных отличительных признаков млекопитающих. У предков зверей — зверозубых рептилий — была только одна слуховая косточка (стремечко), а молоточек и наковальня составляли единое целое с нижней челюстью. То же самое наблюдается и у самых примитивных древних млекопитающих. У современных млекопитающих в ходе эмбрионального развития эти косточки сначала, как и у далеких предков, прилегают к нижней челюсти, а затем отделяются от нее. Отделение происходит в два этапа: сначала слуховые косточки отделяются от боковой поверхности нижней челюсти, но остаются связаны с ней спереди посредством меккелева хряща (этот хрящ, кстати, представляет собой остаток первичной, изначальной нижней челюсти позвоночных; у акул вся нижняя челюсть образована меккелевым хрящом). На втором этапе исчезает и это переднее соединение: меккелев хрящ у взрослых млекопитающих рассасывается. На основе этих фактов более 30 лет назад была выдвинута гипотеза, согласно которой в эволюции среднего уха млекопитающих имелась промежуточная стадия — примерно такая же, как в эмбриональном развитии современных зверей (Allin Е. F. Evolution of the mammalian middle ear. // J. Morphol.1975. 147, 403-438.). Яноконодон блестяще подтвердил эту догадку. Именно поэтому первооткрыватели присвоили ему видовое название allini — в честь Эдгара Аллина (Edgar Allin), автора подтвердившейся гипотезы. Молоточек и наковальня у яноконодона уже отделены от нижней челюсти с боков, но еще прикреплены к ней спереди посредством окостеневшего меккелева хряща. Строение скелета яноконодона еще раз подтвердило, что в ранней эволюции млекопитающих многие прогрессивные признаки появлялись параллельно и независимо в разных линиях. Например, у яноконодона имеются ребра на поясничных позвонках (это примитивный признак), тогда как у некоторых близких форм поясничные ребра отсутствуют. Сравнительно-анатомический и филогенетический (эволюционный) анализ мезозойских млекопитающих показывает, что поясничные ребра многократно исчезали и вновь появлялись в разных эволюционных линиях. По мнению исследователей, в случае с поясничными ребрами, как и с другими признаками, затрагивающими дифференцировку отделов позвоночника, многочисленные параллелизмы могут объясняться тем, что эти признаки контролируются очень небольшим количеством генов. Речь идет о знаменитых Нох-генах, многофункциональных регуляторах развития, которые задают общий план строения, симметрию и дифференцировку отделов тела у многоклеточных животных. Например, если у мыши отключить ген Нох10 (который, правда, присутствует в геноме в трех копиях, и отключить надо все три), то на поясничном отделе позвоночника развиваются ребра. Скорее всего, многократное появление и исчезновение поясничных ребер у древних млекопитающих объясняется простотой "кодирования" этого признака, то есть тем, что для его изменения достаточно всего одной или немногих мутаций.
Yanoconodon allini. Сравнительно простые, эффективные и "умные" системы регуляции индивидуального развития, сложившиеся в ходе эволюции у животных (в том числе система Нох-генов), обладают важным побочным свойством: их случайные изменения (например, в результате мутаций) с довольно высокой вероятностью могут породить не только нежизнеспособных монстров, но и что-то вполне осмысленное и даже удачное. В этом, возможно, состоит одна из причин ускорения прогрессивной эволюции у высших животных (Сходные идеи развивал И. И. Шмальгаузен В книге Факторы эволюции. http://evolbiol.ru/factory.htm. Хотя, разумеется, о Hox-генах в то время еще никто не знал). (Источник: Zhe-Xi Luo, Peiji Chen, Gang Li, Meng Chen. A new eutriconodont mammal and evolutionary development in early mammals // Nature. 2007. V. 446. P. 288-293.)
Параллельная эволюция признака "наличие/отсутствие поясничных ребер " у мезозойских млекопитающих. Генетические основы параллельной эволюции изучены пока слабо. Сходные изменения морфологии или биохимии организмов могут быть обусловлены как одинаковыми, так и совершенно разными наборами мутаций. Какой из этих двух вариантов встречается чаще, до сих пор не известно, и число детально исследованных случаев невелико. В главе "Рождение сложности" мы говорили о том, как бактерии приспосабливаются к антибиотикам. Такое приспособление происходит у разных бактерий параллельно и, по-видимому, может обеспечиваться одним и тем же комплексом мутаций, приобретаемых в определенной последовательности. Однако это крайне простой и редкий случай, когда между признаком (устойчивость к антибиотику) и геном (в данном случае это ген фермента бета-лактамазы) наблюдается соотношение один к одному. Гораздо чаще, особенно у организмов, более сложных, чем бактерии, каждый ген влияет на множество признаков, а каждый признак определяется множеством генов. К тому же связи между генами и признаками опосредуются многочисленными внешними и внутренними факторами в ходе индивидуального развития. Все это делает задачу "расшифровки" генетических основ эволюционных преобразований чрезвычайно сложной. Тонкая подстройка многофункционального гена может приводить к параллельному появлению новых признаков. Группе исследователей из США удалось расшифровать генетический механизм параллельной эволюции пятен на крыльях у самцов мушек-дрозофил. Эти пятна имеются у одних видов рода Drosophila и отсутствуют у других. В процессе эволюции пятна появлялись и утрачивались неоднократно, хотя выглядят они у всех видов примерно одинаково. Исследователи показали, что наличие или отсутствие пятен определяется многофункциональным геном yellow ("желтый"), от которого зависит окраска не только крыльев, но и прочих частей тела. При этом пятнистость крыльев определяется не "основным" (кодирующим) участком гена, в котором записано строение соответствующего белка. Как выяснилось, все дело в том, в каких частях развивающегося крыла происходит экспрессия (работа) гена yellow на стадии куколки. У видов без пятен слабая экспрессия наблюдается равномерно по всему крылу, резко усилена на концах крыльев (как раз там, где у взрослой мухи будут темные пятна).
В ходе эволюции пятна на крыльях дрозофилы несколько раз утрачивались и появлялись – в зависимости от характера экспрессии многофункционального гена yellow. Характер экспрессии гена yellow определяется двумя некодирующими регуляторными участками гена. Пятна на крыльях у дрозофил иногда появлялись благодаря модификации первого из этих участков, иногда — второго. Таким образом, в данном случае независимое появление одного и того же признака (пятен на крыльях) было обеспечено разными генетическими изменениями, затронувшими, правда, один и тот же ген. Исследование показало, что тонкая "подстройка" регуляторных участков многофункциональных генов может приводить к появлению новых морфологических признаков у таких сложных организмов, как дрозофила. (Источник: Benjamin Prudhomme, Nicolas Gompel, Antonis Rokas, Victoria A. Kassner, Thomas M. Williams, Shu-Dan Yeh, John R. True, Ssan B. Carroll. Repeated morphological evolution through cis-regulatory changes in a pleiotropicgene // Nature. 2006. V. 440. P. 1050-1053.). По-видимому, именно изменения регуляторных участков генов играют основную роль в эволюции формы и строения сложных организмов. Большинство генов у таких организмов — многофункциональные. Изменение кодирующей части многофункционального гена может вызвать целый каскад разнообразных малопредсказуемых последствий. Гораздо удобнее и безопаснее чуть-чуть "подкрутить" один из многочисленных регуляторов, влияющий не на все функции данного гена сразу, а лишь на одну или несколько. Важнейший для всей человеческой цивилизации пример параллельной эволюции дают культурные злаки. Ведь все множество одомашненных видов и сортов злаков, от кукурузы до пшеницы, должны были обрести совершенно определенный набор хозяйственных признаков. Это скороспелость, высокая продуктивность, неопадающие семена, голозерность и др. Скороспелость, то есть превращение многолетников или двухлетников в однолетники, необходима, так как урожай желательно иметь каждый год, а не через два года на третий. Неопадающие семена и неломкий колос нужны для предотвращения опадения на землю спелых семян до сбора их земледельцем. Голозерность сильно облегчает и ускоряет процесс обмолота. И все эти признаки культурные растения приобрели в результате неосознанного искусственного отбора независимо друг от друга. Такая параллельная эволюция, по-видимому, сильно облегчалась тем, что формирование многих хозяйственных признаков контролируется единичными регуляторными генами. И если этот единственный регуляторный ген отключить или изменить его работу мутацией, то сразу получится нужный признак. По-видимому, именно таким способом сформировались у многих злаков неопадающие семена и неломкие стебли. Для появления подобного признака часто бывает достаточно "испортить" всего один регуляторный ген из семейства MADS-генов. MADS-тены — это гены растений, аналогичные по функциям Нох-генам животных. Эти гены у культурных растений имеют очень большое сходство, поэтому легко можно ожидать и появления сходных мутаций у разных растений (Н. П. Гончаров и др. Доместикация злаков Старого Света: поиск новых подходов для решения старой проблемы. // Журнал общей биологии. 2007. Т. 68. № 2. С. 126-148. http://elementy.ru/genbio/synopsisPartid=98.). Закономерная повторяемость в неповторимом живом многообразииВ 1920 году на съезде селекционеров в Саратове Н. И. Вавилов сообщил об открытии удивительной закономерности, которой подчиняется наследственная изменчивость у разных видов и родов растений. Оказалось, что вариации в строении листьев, корней, колосьев и семян, наблюдаемые у особей любого вида, вовсе не случайны и не беспорядочны. Каждый вид имеет строго определенный набор возможных вариаций, причем эти наборы ("ряды") почти идентичны у близкородственных видов и очень похожи у представителей разных родов и даже семейств. Изменчивость разных видов злаков можно изобразить в виде таблицы, где каждый ряд соответствует виду, а каждая колонка — определенной вариации. На основе составленной им таблицы Вавилов сформулировал "закон гомологических рядов в наследственной изменчивости". Самое удивительное, что закон гомологических рядов позволил даже предсказать существование таких разновидностей, которые еще не были известны науке! Точно так же, как пустые клетки менделеевской таблицы впоследствии заполнились новооткрытыми химическими элементами, свойства которых в точности совпали с предсказанными, так и пустые клетки вавиловской таблицы были заполнены открытыми природными разновидностями злаков. Главное — знать, что ищешь. Это блестящее подтверждение пришло позже, однако уже после первого доклада Вавилова на съезде селекционеров присутствующие устроили овацию, а один известный ботаник сказал: "Это биологи приветствуют своего Менделеева!" Таким образом, наследственная изменчивость, составляющая основу естественного отбора и эволюции, оказалась строго упорядоченной, а вовсе не "случайной". Такой вывод с большим трудом укладывался в схемы классического дарвинизма. Ведь они основывались на отборе чисто случайных вариаций. Некоторые отечественные биологи-теоретики, такие как А. А. Любищев и С. В. Мейен, придавали вавиловским гомологическим рядам огромное значение и считали, что за этим явлением скрывается некий фундаментальный закон, управляющий, возможно, не только биологической эволюцией, но и всем мирозданием. Они подчеркивали, что гомологические ряды, подобные рядам Вавилова, наблюдаются и в строении кристаллов, и в структуре органических молекул. Интересно и то, что многие признаки, изменчивость которых подчиняется вавиловскому закону, например, сходные вариации формы листьев у очень далеких друг от друга растений — цветковых, голосеменных и папоротников, не имеют явного приспособительного значения. Поэтому происхождение такого сходства трудно объяснить естественным отбором. По-видимому, в вавиловских рядах действительно проявляются некие общие законы развития (самоорганизации) сложных систем, причем не только биологических. Система, состоящая из взаимосвязанных элементов (блоков), может существовать в различных устойчивых состояниях, переходя из одного в другое, но число этих состояний ограничено, а их характер определяется свойствами элементов и структурой их взаимодействий. Очевидно, что дискретный и упорядоченный характер изменчивости накладывает жесткие ограничения на эволюционные возможности организмов. Поэтому существование сходных вариаций у разных видов во многом объясняет то явление, которому посвящена эта глава, — параллельную эволюцию. Сходная изменчивость предопределяет сходство эволюционных изменений у разных групп, а ограниченное число возможных вариаций ограничивает и число возможных путей эволюционного развития. Чем же определяется дискретный характер изменчивости? Прежде всего нужно хорошо понимать, какая это сложная штука — жизнь. Чтобы выжить, любой организм должен приспосабливаться к огромному числу всевозможных внешних факторов. Известно, что никакую систему нельзя оптимизировать одновременно по многим параметрам. Неизбежно приходится что-то выбирать и чем-то жертвовать. Кроме того, организм — это целостная система, вся части которой должны работать согласованно, не мешать, а помогать друг другу. Приспособление к какому-то внешнему фактору одного органа неизбежно влияет на всю систему, так что другим органам уже приходится приспосабливаться не только к выполнению своих собственных задач, но и к произошедшему в самом организме изменению. Иначе вся сложнейшая конструкция может рассыпаться. Но и это еще не все. Каждый многоклеточный организм развивается из одной клетки — яйца. Значит, нужно разработать такую совершенную генетическую программу развития, чтобы в результате последовательных делений одной-единственной клетки в итоге получился сложный жизнеспособный организм. А ведь еще необходимо следить за тем, чтобы жизнеспособность (то есть целостность системы) неизменно сохранялась на каждом этапе роста и развития. Соблюсти все эти условия чрезвычайно сложно. Не случайно некоторые биологи сравнивают эволюцию с переделыванием самолета на лету. Поэтому каждое живое существо — это результат труднейшего компромисса между многими тысячами разнообразных требований, предъявляемых к системе как извне, так и изнутри. Идея о "компромиссном" характере организации живых существ ("метафора адаптивного компромисса" (А. П. Расницын. Процесс эволюции и методология систематики. 2002. http://www.palaeoentomolog.ru/Publ/Rasn/methodology.html.)), помогает понять многое и в строении живых существ, и в их эволюции (см. главу "Рождение сложности"). Во-первых, становится ясно, почему далеко не о каждом признаке и свойстве живого существа можно уверенно сказать, "зачем это нужно". А ведь отсутствие явного адаптивного, то есть приспособительного, смысла у многих признаков всегда смущало дарвинистов. Обычно отговаривались тем, что, дескать, пока мы не знаем, зачем этот признак, но когда-нибудь обязательно узнаем. Ну а теперь становится понятным, что многие признаки появились не "зачем", а "потому что" — это побочные следствия компромисса. Если бы бабочки умели говорить, то на вопрос "почему у тебя на крыльях именно такой узор, а не другой?" они могли бы с полным правом ответить: "Так получилось".
Почему бабочки похожи на цветы? Многих биологов-теоретиков приводил в замешательство "детский" вопрос: почему дневные бабочки, питающиеся нектаром цветов, сами похожи на цветы, а иные еще и издают "цветочный" аромат? Не в деталях, не до полного сходства — это не маскировка, — но в общем и целом, по яркости и броскости рисунка крыльев-лепестков? Мне кажется, здесь проявился "принцип зацепки": естественному отбору, чтобы создать адаптацию, нужно зацепиться за какое-то уже существующее свойство — "преадаптацию". Яркая и необычная окраска бабочек нужна им для привлечения половых партнеров. Бабочки питаются нектаром, и поэтому их органы чувств и нервная система издавна были "настроены" на поиск цветов (и сами цветы, чтобы привлечь опылителей, становились в процессе эволюции более яркими и броскими). Совершенствуя способы привлечения партнера, естественный отбор "зацепился" за эту психологическую особенность бабочек. Преимущество в размножении стали получать те особи, которые привлекали взгляд своих сородичей, порхающих в поисках яркого цветка. Во-вторых, понятно, что достигнутый с таким трудом компромисс — сложнейшую систему индивидуального развития и строения организма — довольно трудно изменить. Ведь в ней все взаимосвязано, изменишь одну часть — и все может развалиться. Пути для изменения все-таки остаются, но их не может быть много, и при этом они дискретны — структура сложной системы предусматривает возможность лишь ограниченного и строго определенного набора изменений. Возможность изменения сохраняется благодаря тому, что живые системы никогда не становятся абсолютно целостными, никогда не сливаются в единый и неделимый "атом". На всех уровнях сохраняется блочный принцип "сборки" живых систем, причем отдельные блоки (гены, клетки, органы, ткани) стараются сохранить хотя бы частичную независимость от других. Благодаря этому далеко не каждое изменение одного блока сразу разрушает систему. Например, в индивидуальном развитии отдельные процессы роста и дифференцировки клеток хоть и сильно взаимосвязаны, но все-таки не абсолютно "слиты" в единую жесткую последовательность. Многие из этих процессов могут меняться независимо от остальных. Например, все земноводные проходят в своем развитии стадию водной личинки. У лягушек это головастики, очень сильно отличающиеся от взрослых лягушек и по строению, и по образу жизни. У хвостатых земноводных — тритонов и саламандр — личинки отличаются от взрослых гораздо меньше. У них другие размеры, плавательная складка на хвосте и спине, ветвистые наружные жабры, которые у взрослых рассасываются, а также ряд второстепенных отличий. Превращением личинки во взрослое животное управляет гормон тироксин. Что же произойдет, если в результате мутации тироксин в организме не образуется или ткани вдруг станут нечувствительны к нему? И то и другое нарушение может произойти в результате одной-единственной "точечной" мутации в одном из генов. Головастик лягушки просто-напросто умрет, не оставив потомства. А вот личинки некоторых саламандр в этом случае не умирают. Все процессы роста и развития у них продолжаются как ни в чем не бывало, за исключением того, что они так и не теряют жабры и плавник и не выходят на сушу. В положенное время у таких личинок развиваются половые железы, и они совершенно нормально размножаются и откладывают икру, из которой выводится потомство, унаследовавшее исходную мутацию. В итоге из наземной саламандры получается чисто водное животное — уже известный нам аксолотль, размножающаяся личинка-переросток. У аксолотля отключилась система превращения личинки во взрослое животное. Все остальные системы продолжают работать по-прежнему. Так что изменение отдельных "блоков" системы индивидуального развития не обязательно приводит к гибели и может даже в некоторых случаях оказаться полезным — например, если у данного вида земноводных на суше больше врагов, чем в воде. Это явление — переход к размножению на ранней стадии, приводящий к сохранению у взрослых особей эмбриональных и личиночных черт, — играло важную роль в эволюции. Возможно, таким путем появились позвоночные из головастико-подобных личинок сидячих морских фильтраторов — оболочников (Tunicata). Есть теория, что и в происхождении человека какую-то роль сыграли подобные процессы. По ряду признаков, таких как безволосость и высокий лоб, человек больше похож на эмбрионов и детенышей обезьян, чем на взрослых. Другая возможность скачкообразных перестроек состоит в перенесении свойств одних частей тела на другие. Как мы знаем, для этого может быть достаточно мутации одного гена-регулятора. В результате какой-нибудь "переключатель", управляющий дифференцировкой частей организма, вдруг начинает включаться не в то время или не в том месте. Так, у различных насекомых обнаружены вполне "вавиловские" гомологичные ряды мутаций, приводящих к тому, что свойства одних сегментов тела или конечностей проявляются в других сегментах, неположенных. Вместо усиков могут вырасти ноги, вместо третьего сегмента груди развивается точная копия второго, и если это произойдет у мухи, то она в результате автоматически приобретет вторую пару крыльев! Или же ноги вырастут не только на грудных, но и на брюшных сегментах, и получится насекомое с восемьюдесятью парами ног. Правда, такие мутанты обычно умирают в раннем возрасте. Мутации такого типа — их называют "гомеозисными" — могли сыграть большую роль в эволюции. В результате гомеозисной мутации животное, подобное кембрийскому аномалокарису с единственной парой членистых конечностей, могло бы приобрести сразу множество таких ног вдоль всего тела. Среди древних ископаемых рыб силурийского и девонского периодов есть группы, имеющие только одну пару плавников — переднюю. Но известны и такие ископаемые рыбы, у которых множество парных плавников. Все современные рыбы, как и наземные позвоночные, имеют две пары конечностей. Я нисколько не удивлюсь, если в скором времени генетики докажут, что изменения числа пар конечностей в ранней эволюции позвоночных происходили за счет гомеозисных мутаций, затрагивающих Нох-гены. По предположению академика JI. П. Татаринова, шерстяной покров у млекопитающих тоже мог появиться в результате гомеозисной мутации. У предков млекопитающих — звероподобных рептилий — некоторые чешуи на морде удлинились и стали выполнять осязательную функцию. Постепенно они превратились в длинные упругие волоски — вибриссы. А затем в один прекрасный день в результате гомеозисной мутации все остальные чешуи на теле превратились в такие волоски. Правда, чувствительной функции у них при этом не появилось, поскольку к ним не подходили соответствующие нервы (Точно так же обстоит дело и с ногами насекомых, вырастающими на месте усиков в результате гомеозисных мутаций: нервы, необходимые для работы ног, к ним не подходят, а те нервы, что управляют работой усиков, "не годятся", поэтому лишние ноги остаются неподвижными, несмотря на нормально развитую мускулатуру), но для утепления мутация оказалась очень даже полезной. Можно подумать, что подобные мутации не могут привести к усложнению организмов, потому что это не более чем перетасовка и перекомбинация имеющихся признаков. Это, однако, не совсем так. Попадая в новое место, новое клеточное и тканевое "окружение", перепрыгнувший орган неизбежно и сам становится немножко другим. Так закладывается основа для дифференцировки. Например, пресловутые ноги, вырастающие иногда у насекомых вместо усиков, на самом деле довольно сильно отличаются от обычных ног и могли бы стать основой для появления совершенно нового типа конечностей, если бы... нормальные усики на этом месте не были так необходимы этим глупым насекомым. Постепенно или скачками?Из дискретного характера изменчивости, казалось бы, следует, что эволюционные преобразования в основном должны происходить внезапными скачками. Однако, как и любые другие эволюционные закономерности, закон гомологических рядов Вавилова, правило "квантованной" изменчивости и скачкообразных переходов живых систем из одного устойчивого состояния в другое не нужно абсолютизировать. Далеко не всякая изменчивость дискретна, и многие эволюционные преобразования организмов происходят вполне плавно.
Градуализм и сальтационизм. Вопрос о равномерности темпов эволюции довольно долго был одной из самых острых проблем в эволюционном учении. И сам Дарвин, и многие его последователи видели эволюцию как процесс в основном плавный, постепенный, то есть градуалистический. Высказывалась, правда, и другая точка зрения — о возможности мгновенного превращения одного вида в другой в результате крупномасштабных мутаций, или, по-другому, макромутаций. Это учение называют сальтационизмом, от "сальто" — прыжок. Основным аргументом противников сальтационизма было то, что крупные мутации почти всегда смертельны или по крайней мере очень вредны. Сальтационисты на это отвечали, что среди "монстров", появляющихся в результате макромутаций, могут оказаться и "перспективные" (hopeful monsters). Открытие Нох-генов и гомеозисных мутаций показало, что в фантазиях сальтационистов было разумное зерно. Однако классическая СГЭ образца 50-х - 60-х годов прошлого века придавала основное значение отбору небольших наследственных отклонений и считала эволюцию в основном постепенным, или градуалистическим, процессом. Единственным источником наследственной изменчивости сторонники СТЭ считали случайные мутации — ошибки при копировании ДНК. Считалось, что, поскольку мутационный процесс совершенно случаен и не направлен, то и идти он должен примерно с одинаковой скоростью у всех живых организмов. Это предположение легло в основу принципа "молекулярных часов", который активно используется и по сей день. На основе этого принципа исследователи по числу различий в нуклеотидных последовательностях ДНК оценивают время расхождения видов, то есть определяют, когда жил их последний общий предок. И метод молекулярных часов обычно не так уж плохо работает, что само по себе может показаться странным. Ведь сегодня хорошо известно, что мутагенез не всегда является абсолютно случайным, его темп может целенаправленно регулироваться клеткой, в разных группах организмов и даже в разных частях одного и того же генома он идет с разной скоростью. Чтобы молекулярные часы продолжали тикать, приходится добавлять ограничения, выбирать "подходящие" куски генома, учитывать особенности группы, принимать в расчет разницу между значимыми и "молчащими" нуклеотидными заменами. Альтернативой градуализму (учению о постепенной эволюции) и сальтационизму (эволюции путем скачков) стал пунктуализм, или теория прерывистого равновесия, сформулированная в 1972 году Стивеном Гулдом и Нильсом Элдриджем. Эта концепция предполагает, что в эволюции видов чередуются длительные периоды стабильности, когда основные черты вида сохраняются неизменными, и короткие периоды быстрых изменений, в ходе которых вид преобразуется. Он либо целиком превращается в другой вид, либо делится на два или более новых вида, либо "отпочковывает" их от себя. Точка зрения пунктуалистов, основанная на обширном палеонтологическом материале, имеет и достаточно веские теоретические обоснования. В самых общих чертах их можно свести к следующему. Вид представляет собой относительно устойчивую самоподдерживающуюся систему. Чтобы на месте старой системы возникла новая, старая должна быть разрушена. Нужна хорошая "встряска", приводящая к разрушению внутренних связей. В популяциях живых организмов разрушение старой системы связей, приводящее к дестабилизации системы, проявляется прежде всего в резком росте изменчивости. Имеются экспериментальные подтверждения того, что резкое изменение условий приводит не к плавному и постепенному сдвигу морфологических характеристик популяции, а к быстрому росту изменчивости, "размыванию" признаков, и только потом из этой дестабилизированной популяции может выкристаллизоваться новая разновидность или вид (См.: А. С. Раутиан. Правило дестабилизации, http://www.evolbiol.ru/rautian.htm#destabilization.). Поначалу споры градуалистов с пунктуалистами были довольно бурными, но сегодня эти две концепции вполне мирно уживаются друг с другом. Их справедливо считают не противоречащими друг другу, а взаимодополнительными. Обе идеи быстро обрастают всевозможными примерами, уточнениями и обоснованиями. Имеющиеся факты говорят о том, что эволюция иногда идет по пунктуалистическому, иногда — по градуалистическому сценарию. Обе концепции, однако, до сих пор относятся к числу "эмпирических обобщений", а не строгих теорий — как, впрочем, и почти все остальные так называемые "законы эволюции". В немалой степени это объясняется тем, что никому пока не удалось выяснить точное количественное соотношение градуалистической и пунктуалистической составляющих в эволюционном процессе. Доказан скачкообразный характер эволюционного процесса. В 2006 году британские биологи предприняли серьезную попытку решить давний вопрос о соотношении постепенности и скачкообразности в эволюции. Методика, примененная исследователями, проста, как все гениальное. Странно, что никто до этого не додумался раньше. Суть идеи в том, что искомое соотношение легко можно вычислить, сопоставив между собой длины ветвей эволюционных "деревьев", которые строятся при помощи стандартных методик на основе величины генетических различий между сравниваемыми видами. Ветви таких деревьев имеют определенную длину, соответствующую величине генетических различий или, что то же самое, величине эволюционных изменений, произошедших в данной эволюционной линии. Все виды, используемые в таком анализе, обычно современные (выделение ДНК из ископаемых остатков — это все-таки экзотика). Значит, если эволюция шла равномерно (градуалистически), то расстояние от основания древа до кончика любой из ветвей (эти кончики соответствуют анализируемым видам) должно быть одинаковым, и оно не должно зависеть от количества "узлов" (точек ветвления), расположенных между кончиком ветви и корнем. Если же эволюция ускорялась в момент ветвления, то есть шла по пунктуалистическому сценарию, то расстояние от корня до конца ветви должно быть пропорционально числу узлов, расположенных между ними (см. рисунок). Исследователи проанализировали эволюционные деревья, построенные по молекулярным данным для 122 групп близкородственных видов. В 57 случаях из 122 между обнаружилась вполне четкая, статистически достоверная корреляция между длиной ветви и числу узлов на ней. Для остальных деревьев корреляция оказалась недостоверной — в основном по той причине, что у этих деревьев было слишком мало ветвей, то есть количество видов в выборке оказалось недостаточным для получения статистически надежных результатов.
Если эволюция идет в соответствии с принципом "прерывистого равновесия", то есть скачками, эволюционные деревья должны выглядеть как на рисунке А. Если же эволюция идет с постоянной скоростью, правильным окажется рисунок В. Таким образом, пунктуалистический эффект действительно существует и проявляется если и не всегда, то достаточно часто. Любопытно, что у растений и грибов он проявляется заметно сильнее, чем у животных. Исследователи не остановились на достигнутом и определили также относительный вклад пунктуалистического эффекта в суммарную величину различий между нуклеотидными последовательностями ДНК разных видов. Выяснилось, что в среднем около 22% наблюдаемых различий возникает во время "взрывных" периодов видообразования, а остальные 78% постепенно накапливаются в ходе градуалистической эволюции. В работе рассматривались только различия в нуклеотидных последовательностях, причем функциональное значение этих различий никак не учитывалось (да этого никогда и не делают при построении "молекулярных деревьев"). На уровне организма картина может быть иной. Известно, что значительная часть изменений ДНК никак не отражается на строении организма, и, с другой стороны, даже очень небольшие изменения в ключевых участках ДНК могут привести к радикальным изменениям организма. Вполне возможно, что те изменения ДНК, которые происходят очень быстро в период видообразования, связаны с более значительными изменениями в строении организма, чем те, что происходят во время градуалистического этапа эволюции. В этом случае в морфологической эволюции будет значительно больше "пунктуализма", чем в эволюции молекулярной. Что, собственно, и наблюдается в палеонтологической летописи. Данная работа интересна как еще одна попытка строгого количественного обоснования одного из эмпирических "законов эволюции". Все-таки приятно, что давно подмеченные зоологами и палеонтологами, но не доказанные закономерности эволюции постепенно начинают получать строгие обоснования. (Источник: Mark Pagel, Chris Venditti, Andrew Meade. Large Punctuational Contribution of Speciation to Evolutionary Divergence at the Molecular Level // Science. 2006. V. 314. P. 119-121.) Иногда кажущаяся дискретность изменений порождается отбором, который благоприятствует некоторым из возможных вариантов изменений, но далеко не всем. Например, когда организм человека вырабатывает иммунитет против новой инфекции, в генах защитных белков — антител — происходит активное мутирование. Это необходимо, чтобы "методом случайного поиска" подобрать такое антитело, которое будет лучше всего узнавать нового возбудителя и прикрепляться к нему (подробнее об этом см. в главе "Управляемые мутации"). Сравнивая структуру получающихся при этом антител, ученые обнаружили, что больше всего мутаций происходит в трех небольших участках так называемой вариабельной области (V-области) гена, кодирующего антитело. Сложнейшими экспериментами было доказано, что мутированию подвергаются не только эти участки, а вся V-область, причем мутации везде происходят с одинаковой скоростью. Почему же в имеющихся в крови антителах измененными обычно оказываются только три маленьких участка? Оказалось, что это результат отбора. Каждое новое антитело испытывается орагнизмом, и все не подходящие к данному возбудителю антитела отбраковываются. Только мутации, произошедшие в трех небольших участках гена, могут улучшить связывание возбудителя. Если эффективное антитело успешно прошло проверку, то только тогда оно начинает производиться в больших количествах. А мутации других участков почти всегда вредны, и такие антитела организм отбраковывает. Понятно, что отбор такого рода может отчасти объяснять и происхождение некоторых вавиловских рядов. Тем не менее остается в силе общее правило, гласящее, что структура организма допускает лишь ограниченный набор возможных преобразований и тем самым придает эволюции некоторую направленность, а иногда и предопределенность. По-видимому, эволюция — это процесс в целом закономерный и предопределенный, но в деталях и частностях случайный. Предсказать ход эволюции можно лишь в самых общих чертах. У эволюции нет строгих законов, как в математике или физике. У нее есть только набор закономерностей и правил, каждое из которых имеет множество исключений. К числу важнейших закономерностей эволюции я бы отнес следующие три "правила". 1. Общая направленность от простого к сложному. Хотя до сих пор сохранились и процветают такие примитивные формы жизни, как бактерии, никто не станет отрицать, что в биосфере идет постепенное появление и накопление все более сложных организмов. Часто усложнение организации оказывается выгодным, потому что ведет к повышению интенсивности обмена веществ (росту "энергии жизнедеятельности"), а ведь в биологической эволюции, как и в химической (см. главу "Происхождение жизни"), побеждает тот "круговорот", который крутится быстрее. Это позволяет более сложным организмам занимать господствующее положение в сообществах, оттесняя примитивных предков в менее привлекательные ниши. Именно поэтому облик биосферы определяется в основном высокоорганизованными животными и растениями. И это несмотря на то, что и по численности, и по массе бактерии их явно превосходят. Рост биоразнообразия в целом носит аддитивный, то есть накопительный, характер — новое обычно добавляется к старому, а не вытесняет его. "Новое не вытесняет старое, а добавляется к нему". Динамика разнообразия трех крупнейших групп наземных растений: споровых (Pteridophyta) — это самая древняя и примитивная группа; голосеменных (Gymnospermophyta); покрытосеменных,, или цветковых (Magnoliophyta), — это самая молодая и прогрессивная группа. По горизонтальной оси — геологическое время (млн лет назад). Видно, что развитие новых "прогрессивных" групп не сопровождалось существенным сокращением разнообразия (вытеснением) их более примитивных предков. Из статьи: Марков А. В., Коротаев А. В. Гиперболический рост разнообразия морской и континентальной биот фанерозоя и эволюция сообществ. // Журнал общей биологии. 2008. № 3. 2. Рост устойчивости и приспособляемости живых систем. Все эволюционные линии, дожившие до наших дней, в ходе своего развития прошли через горнило множества экологических кризисов, катастроф и массовых вымираний. Те группы, которые не могли быстро приспосабливаться к меняющимся условиям, в большинстве своем давно вымерли. Устойчивые, пластичные линии постепенно накапливались в биосфере. Это видно, например, из того факта, что с течением времени средняя продолжительность существования видов, родов и семейств неуклонно росла. Поэтому в наши дни биосферу населяют самые устойчивые и пластичные формы жизни из всех когда-либо существовавших.
Рост средней продолжительности существования (ПС) семейств в течение фанерозойского зона (542 млн лет назад — наши дни). Тонкая линия — семейства морских организмов, толстая — континентальных. Различия в колебаниях двух кривых в данном случае не столь важны (известноу что в океане и на суше эволюция шла во многом по-разному). Важнее, что оба графика являются возрастающими. По вертикальной оси: средняя ПС семейств в млн лет. По горизонтальной оси — геологическое время (млн лет назад). Из статьи: Марков А. В., Коротаев А. В. Гиперболический рост разнообразия морской и континентальной биот фанерозоя и эволюция сообществ. // Журнал общей биологии. 2008. № 3. 3. Рост эффективности и безотходности биогеохимического круговорота. С ростом сложности и совершенства организмов и их сообществ неизбежно растет и эффективность глобального круговорота веществ, в котором биосфера играет важнейшую роль и который определяет "лицо" нашей планеты. Например, самое сложное и совершенное из современных сообществ — тропический лес — не только чрезвычайно быстро "прокручивает" через себя огромные количества вещества и энергии, но и практически не производит никаких отходов. Там не образуется даже подстилка из листьев и других отмерших частей растений — все это очень быстро перерабатывается грибами, бактериями и беспозвоночными и возвращается в круговорот. Совсем по-другому обстояло дело, например, в древних лесах каменноугольного периода, в которых из-за несовершенства структуры сообщества огромные массы отмершей древесины накапливались, образуя месторождения каменного угля. В результате столь необходимый для жизни углерод безвозвратно выводился из глобального круговорота (Человек, сжигая каменный уголь, снова возвращает этот "потерянный" углерод в биосферу и делает его доступным для живых организмов. При сжигании угля образуется углекислый газ, из которого растения создают органические вещества в процессе фотосинтеза). Рост безотходности заметен и в эволюции организмов. У высших растений и животных постепенно растет продолжительность жизни, снижается "детская смертность", развивается забота о потомстве, что позволяет, в свою очередь, снизить уровень рождаемости — то есть фактически производить меньше заведомо обреченных на гибель потомков. Все три названных закономерности: усложнение, рост устойчивости и безотходности — отчетливо прослеживаются и в развитии человеческого общества. Это позволяет говорить о преемственности социальной эволюции по отношению к эволюции биологической и придает особый смысл и практическое значение эволюционным исследованиям. Важно подчеркнуть, что из этой преемственности вовсе не следует никакого "социал-дарвинизма" и она вовсе не свидетельствует об усилении "борьбы за существование" и "естественного отбора" в обществе, как пытались доказать некоторые политические силы в первой половине ХХ века. В трудах современных эволюционистов, например, В. А. Красилова, подчеркивается неуклонное ослабление роли борьбы за существование и отбора в ходе эволюции, развитие более "гуманных" эволюционных стратегий, основанных на взаимопомощи и симбиозе и ведущих к росту пластичности и снижению всевозможных потерь и отходов (см. также главу "Великий симбиоз"). Может быть, самый главный из всех эволюционных законов — это постепенное отступление Смерти и Хаоса перед лицом развивающейся Жизни. Что почитать на эту тему в ИнтернетеН. И. АНДРЕЕВ, С. И. АНДРЕЕВА. Эволюционные преобразования двустворчатых моллюсков Аральского моря в условиях экологического кризиса. 2003. http://evolbiol.ru/andreevy.htm Л. С. БЕРГ. Номогенез, или эволюция на основе закономерностей. 1922. http://rogov.zwz.ru/Macroevolution/berg1922.djvu Н. И. ВАВИЛОВ. Закон гомологических рядов в наследственной изменчивости. Л.: Наука. 1987. http://evolbiol.ru/vavilov.htm В. А. КРАСИЛОВ. Труды по теории эволюции: Нерешенные проблемы теории эволюции. Владивосток: ДВНЦ АН СССР. 1986. http://evolbiol.ru/kr.htm Метаэкология. М.: ПИН РАН. 1997. http://evolbiol.ru/krasilov.htm Е. Н. КУРОЧКИН О четырехкрылом динозавре и происхождении птиц. 2004. // Природа. http://evolbiol.ru/kurochkin2.htm Новые идеи о происхождении и ранней эволюции птиц. 2001. http://evolbiol.ru/kurochkin.htm А. В. МАРКОВ. Обзоры и подборки литературы на сайте "Проблемы эволюции" 2003-2007: Ароморфозы и параллельная эволюция, http://evolbiol.ru/paral/paral.htm Основные этапы прогрессивной эволюции. Важнейшие ароморфозы. http://evolbiol.ru/aro.htm Эволюция на основе случайности или закономерности? http://evolbiol. ru/determinizm.htm Параллелизмы и гомологическая изменчивость: причины и следствия. http://evolbiol.ru/paral_homol.htm С. В. МЕЙЕН. Проблема направленности эволюции. 1975. http://evolbiol.ru/meyen_1975.htm И. Ю. ПОПОВ. Идея направленной эволюции: история и современность. 2006. http://evolbiol.ru/popov.htm А. С. РАУТИАН. Правило дестабилизации. 1988. http://www.evolbiol.ru/rautian.htm#destabilization А. С. СЕВЕРЦОВ." Направленность эволюции. 1990. http://rogov.zwz.ru/Macroevolution/severtsevi990.pdf Дж. СИМПСОН. "Великолепная изоляция" http://rogov.zwz.ru/macroevolution/simpsoni983.djvu А. Н. СОЛОВЬЕВ, А. В. МАРКОВ. Ранние этапы эволюции неправильных морских ежей. 2004. // Экосистемные перестройки и эволюция биосферы. Вып. 6. М.: ПИН РАН, 2004. С. 77-86. http://evolbiol.ru/sbornik04/04markov.htm Л. П. ТАТАРИНОВ. Параллелизмы и направленность в эволюции млекопитающих: Параллелизмы и их эволюционное значение. //Очерки по теории эволюции. М.: Наука. 1987. http://evolbiol.ru/tatparal.htm Необратимость эволюции и ее направленность. // Очерки по теории эволюции. М.: Наука. 1987. http://evolbiol.ru/tatnapr.htm Морфологическая эволюция териодонтов и общие вопросы филогенетики. М.: Наука. 1976. http://evolbiol.ru/tatar.htm И. И. ШМАЛЬГАУЗЕН. Факторы эволюции. Теория стабилизирующего отбора. Издание второе (переработанное и дополненное). М.: Наука. 1968. http://evolbiol.ru/factory.htm |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||