|
||||
|
|
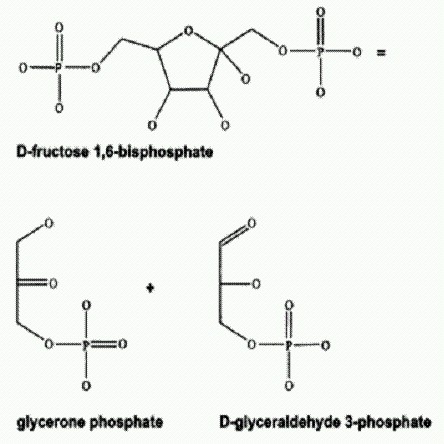
ГЛАВА 4. РОЖДЕНИЕ СЛОЖНОСТИПорядок из хаосаИзлюбленным аргументом креационистов является утверждение о невозможности самопроизвольного развития сложных структур из простых на основе случайных изменений. Действительно, здравый смысл вроде бы подсказывает, что, сколько бы мы ни ворошили лопатой кучу мусора, ничего полезного и принципиально нового из обрывков и обломков само собой не соберется. Креационисты очень любят приводить эту аналогию (она известна во многих вариантах). Часто в этом контексте поминают также второе начало термодинамики, согласно которому якобы невозможен самопроизвольный рост упорядоченности — сам собой нарастать может только хаос. Все это, сказать по правде, полная ерунда. Подобные рассуждения изобилуют логическими ошибками: аналогии не соответствуют объясняемому феномену, а физические законы привлекаются не к месту, то есть делается попытка применить их за пределами установленной для них области применимости. Например, второе начало термодинамики справедливо только для изолированных систем, которые не получают энергии из внешней среды. Живые организмы, напротив, всегда получают энергию извне и, расходуя часть этой энергии на повышение упорядоченности внутри себя, создают при этом порядочный хаос снаружи. Суммарная неупорядоченость (энтропия) всей системы (организм + среда) при этом растет. Поэтому никакого противоречия между развитием жизни и вторым началом термодинамики попросту не существует. Возможность самоорганизации — самопроизвольного рождения порядка из хаоса — показана экспериментально и обоснована теоретически для самых разных типов открытых неравновесных систем. Кроме того, говоря о "случайности", якобы лежащей в основе предполагаемых механизмов эволюции, креационисты занимаются откровенным передергиванием. Они спекулируют на многозначности и расплывчатости термина "случайность". В действительности эволюция основана не на случайностях, а на вполне строгих закономерностях (о которых мы еще будем говорить). Даже мутации, которые до недавнего времени действительно было принято считать случайными, на самом деле далеко не всегда таковы (см. главу "Управляемые мутации"). Дарвиновский механизм естественного отбора сам по себе вполне достаточен для того, чтобы придать эволюционному процессу упорядоченность. Пусть даже первичные изменения (мутации) происходят случайно — благодаря действию отбора запоминание системой произошедших изменений происходит уже не случайно, а строго закономерно. Это избирательное запоминание и производит новую информацию и новую сложность. Между прочим, математикам хорошо известна так называемая S-теорема Ю. Л. Климонтовича, представляющая собой строгое математическое доказательство того, что новая информация (или "отрицательная энтропия", упорядоченность) порождается, сочетанием случайного изменения состояния системы с последующим необходимым (избирательным) запоминанием результатов изменения. Иными словами, эволюционный механизм, предложенный Дарвином, является совершенно адекватным и достаточным объяснением самопроизвольного усложнения живых систем в ходе эволюции — как с точки зрения биологии, так и с точки зрения физики и математики (См. Н.М. Борисов Эволюция, случайность, энтропия (о том, как Дарвин и Клаузиус оказались правыми одновременно) http://evolbiol.ru/evidence10.htm#borisov1). Чтобы убедиться своими глазами в возможности самозарождения сложного из простого, налейте в ванну воды и выньте пробку слива (тем самым превратив закрытую систему в открытую). Вскоре хаотическое движение молекул воды под воздействием силы тяжести на ваших глазах породит довольно сложную, упорядоченную структуру — воронку-водоворот. Когда видишь это впервые в жизни (я помню свое впечатление из раннего детства), возникает ощущение чуда. Почему? Да просто наше мышление так устроено: оно специализировано для целеполагания, мы привыкли планировать свои действия, ориентируясь на ожидаемый результат. Поэтому нам и кажется, что если чего-то не предусмотришь, не проконтролируешь, то ничего хорошего и не выйдет — только разруха и хаос. Раз за разом нас подводит эта "инструментальная" направленность нашего мыслительного процесса. Так, в высокоразвитых сельскохозяйственных цивилизациях Мексиканского нагорья царило убеждение, что если пустить дело на самотек, то ни солнце утром не взойдет, ни лето в положенный срок не настанет, а потому необходимо регулярно подкармливать жертвенной кровью тех светлых богов, что в непрестанной героической борьбе с силами хаоса поддерживают привычный нам миропорядок. Ныне те боги, хочется верить, уже умерли с голоду, но на смене дня и ночи это, как видим, не отразилось. Аналогии с мусорной кучей и лопатой, или с землетрясением, которое может только разрушить здание, но никогда не соберет его из камней, или с мартышкой, которая, случайно нажимая на клавиши, никогда не напишет "Войну и мир", имеют примерно такое же отношение к эволюции, как ритуальная практика ацтеков — к вращению Земли вокруг солнца. Те эволюционные процессы, в результате которых рождается новая сложность, очень мало похожи на эти воображаемые ситуации. Если какое-то сходство и есть, то очень неполное, ограниченное отдельными признаками, поверхностное и ничего не говорящее о сути дела. Мы уже упомянули вскользь об упорядочивающей и направляющей роли отбора и о неслучайности многих мутаций (наследственных изменений). Кроме того, "обрывки и обломки", из которых в процессе эволюции собирается новое, совсем не похожи на те разнородные предметы, которые можно обнаружить в мусорной куче. Начать с того, что материал наследственности (ДНК), генетический код и базовая "технология" его прочтения и реализации у всех организмов одни и те же. Этот факт, между прочим, рассматривается учеными как решающее доказательство происхождения всего живого на нашей планете от единого общего предка (будь то предковый вид или предковое сообщество). Жизненные задачи тоже у всех организмов сходны (выжить, добыть нужные вещества и энергию, вырасти, противостоять внешним разрушающим воздействиям, оставить потомство). Поэтому все имеющиеся в наличии на нашей планете фрагменты ДНК, во-первых, понятны для всего живого (могут быть более-менее адекватно прочтены и "интерпретированы" любой живой клеткой (Это доказывается, в частности, успехами генной инженерии. Гены одних организмов успешно работают в клетках других. Благодаря этому, например, диабетики сегодня обеспечены инсулином — человеческим белком, произведенным бактериями, которым пересадили человеческий ген)), во-вторых, все они являются фрагментами устройств, сходных по своим функциям. Это резко повышает вероятность того, что случайное комбинирование фрагментов (например, в ходе горизонтального обмена генами, о котором рассказано в главе "Наследуются ли приобретенные признаки?") может породить что-то новое и осмысленное. Была раньше такая хорошая игрушка — радиоконструктор (что-то вроде нынешнего "Лего", только из конденсаторов, ламп, катушек и прочих радиодеталей). Простые работающие устройства — такие, например, как детекторный приемник — из этого конструктора собирались с достаточно высокой вероятностью путем абсолютно случайного, произвольного соединения деталек. Этот пример показывает, что если "детальки" хороши, то даже случайное их комбинирование вполне может породить что-то полезное и жизнеспособное. А в "эволюционном конструкторе" детальки, несомненно, самого высшего сорта — отшлифованные естественным отбором, проверенные и испытанные многими поколениями живых "испытателей". Кроме того, геном организма или отдельный ген — это далеко не "Война и мир", а нечто гораздо более помехоустойчивое. Если мы заменим случайным образом букву, слово или предложение в талантливом романе, то наверняка хоть чуть-чуть, но испортим произведение. Если мы заменим в гене нуклеотид или в белке аминокислоту, с очень большой вероятностью не произойдет абсолютно ничего плохого (более того, есть даже вполне реальный шанс, что изменение окажется полезным, но об этом чуть позже). Белковая молекула обычно состоит из нескольких сотен аминокислот, но только очень немногие из них действительно необходимы для того, чтобы белок исправно выполнял свою функцию. Если речь идет о ферменте, то для его работы абсолютно необходимы в основном те аминокислоты, которые составляют так называемый "активный центр". Активный центр — это то место белковой молекулы, которое, собственно, и катализирует реакцию. Кроме того, могут оказаться важными и некоторые аминокислоты, определяющие пространственную конфигурацию молекулы — то, в какую фигуру она самопроизвольно "свернется" после того, как будет синтезирована. Например, в построении пространственной структуры фермента участвуют цистеины — это аминокислоты, содержащие серу, которые образуют дисульфидные мостики (-S-S-), скрепляющие между собой различные витки, лопасти и спирали белковой молекулы. Такие принципиально важные аминокислоты составляют лишь малую часть белка. Более того, это даже не конкретные наборы аминокислот, которые должны находиться в строго определенных местах белковой молекулы, а довольно расплывчатые "рисунки", "паттерны", или, как их официально называют, мотивы. Белки с одинаковой функцией могут сильно различаться по структуре. Вот аминокислотная последовательность фермента фруктозо-бисфосфат альдолазы жгутиконосца Euglena gracilis. Это один из ферментов гликолиза — важного биохимического процесса, в ходе которого, как мы помним, клетка тратит часть энергии, заключенной в молекуле глюкозы, для синтеза двух молекул АТФ без использования кислорода. Каждая буква соответствует определенной аминокислоте (Р — пролин, D — аспарагиновая кислота, F — фенилаланин, К — лизин и т.д.): PDFPKDLKGV LDGNQVRTLF DFAQKKGFAI PAVNCTSSSTVNWLERARD THNPVIIQVS QGGAAFYCGK GVKDEKLIAS VDGSVALAHH VRAVAHTMAP VWHSDHCAK KLLPWFDGML DADGEIFCEH GVPLFSSHML DLSEENDEED IGTCVKYFTR MAKLNLWLEM EIGMTGGVED GVDNSGVAND KLYTSSEQVF AVHKALGASS PNFSIAAAFG NVHGVYKPGN VKLQPNLLKE HQDYARKQLS SSEDHPLYLW FHGPSGSTDA EIHEAVRNGVVKMNLDTDMQ WAYWDGLRQF EAKKHDYLQG QIGNPEGPDK PNKNYYDPRK WIREAELGML ARVKVAFKAV ELPGGLKEFI GIP (Все "расшифрованные" последовательности генов и белков лежат в Интернете в открытом доступе. Данная последовательность доступна по адресу: http://www.ebi.uniprot.org/entry/Q42729_euggr.). А вот тот же самый фермент бактерии Mycoplasma pneumoniae: MLVNIKQMLQ HAKQHHYAVP HININNYEWA KAVLTAAQQA KSPIIVSTSE GALKYISGHQ VWPMVKGLV DALKITVPVA LHLDHGSYEG CKAALQAGFS SIMFDGSHLP FQENFTKSKE LIELAKQTNA SVELEVGTLG GEEDGIVGQG ELANIEECKQIATLKPDALA AGIGNIHGLY PDNWKGLNYE LIEAIAKATN LPLVLHGGSG IPEADVKKAI GLGISKLNIN TECQLAFAKA IREYVEAKKD LDTHNKGYDP RKLLKSPTQA IVDCCLEKMQ LCGSTNKA (http://www.ebi.uniprot.org/entry/alf_mycpn). На первый взгляд заметить сходство между этими белками очень непросто. На второй, впрочем, тоже. А ведь это, по молекулярно-генетическим меркам, весьма похожие друг на друга, близкородственные белки! У них есть длинный сходный участок (от 16-й до 333-й аминокислоты первой молекулы и от 5-й до 265-й аминокислоты второй), в пределах которого совпадает целых 25% аминокислот. Кроме того, некоторые другие аминокислоты в двух белках хотя и разные, но близкие по своим химическим свойствам.
Реакция, которую катализирует фермент фруктозо-бисфосфат альдолазы. Для того чтобы обнаружить это сходство и оценить его количественно, можно воспользоваться специальной программой BLAST, свободно доступной в Интернете по адресу http://www. ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi. TaM надо просто вставить две аминокислотные последовательности в два окошка и нажать кнопку. Такой уровень сходства при сравнении белков представителей разных надцарств живой природы (мы сравнивали эукариот с бактериями) считается очень высоким. В данном случае, возможно, этот результат объясняется тем, что предки эвглены сравнительно недавно приобрели соответствующий ген путем горизонтального переноса от каких-то бактерий. У высших эукариот фруктозо-бисфосфат альдолазы этого семейства вообще не встречаются. У нас с вами, например, они совсем другие. Читатель может попробовать самостоятельно сравнить между собой фруктозо-бисфосфат альдолазы различных организмов (найти их можно, например, в базе данных PFAM http://pfam.sanger.ac.uk/, осуществив поиск по ключевым словам "fructose bisphosphate aldolase"). Любой желающий может легко убедиться, что если сравнить один и тот же (по функции) фермент, взятый у человека и, допустим, у кишечной палочки Escherichia coli, то программа сравнения, скорее всего, выдаст безрадостное "no significant similarity found" — "значимого сходства не обнаружено". Это, впрочем, не значит, что никакого сходства действительно нет — оно есть (это те самые неопределенные "мотивы", о которых шла речь выше), но для его выявления необходимы более мощные аналитические средства, чем программа BLAST. Тем не менее эти столь разные белки примерно с одинаковой эффективностью выполняют одну и ту же работу, то есть катализируют одну и ту же биохимическую реакцию. Конечно, не все различия между разными белками, выполняющими одну и ту же функцию, случайны и бессмысленны. Некоторые из них обеспечивают тонкую подстройку фермента под конкретные особенности той или иной клетки. Однако, как показывает генно-инженерная практика, довольно часто эти белки оказываются вполне взаимозаменимыми. Это значит, что если заменить у какого-либо организма его собственный ген на чужой, кодирующий фермент, тот же по функции, но иной по структуре, то такой генно-модифицированный организм с большой вероятностью окажется вполне жизнеспособным. Из этого видно, что "подобрать" белок с нужной функцией методом случайного перебора аминокислот в принципе не так уж сложно. Для этого вовсе не требуется правильно угадать все несколько сотен аминокислот, составляющих молекулу белка. Достаточно подобрать лишь некий довольно расплывчатый "мотив". Эксперименты показывают, что такая технология отлично работает. Среди коротких белковых молекул, искусственно синтезированных путем случайного комбинирования аминокислот, удается отыскать белки практически с любой каталитической активностью. Конечно, эффективность таких катализаторов ниже, чем у настоящих ферментов, но это неважно. Главное, чтобы отбору было за что "зацепиться", и он со временем доведет функцию до совершенства. Эта технология годится не только для белков, но и для функциональных РНК. Именно таким способом исследователи ищут и находят новые рибозимы — РНК с ферментативными свойствами (см. главу "Происхождение жизни"). Отобрав подходящие молекулы, можно затем раз за разом вносить в них небольшие случайные изменения и отбирать лучшие из получившихся вариантов. Это называют "методом искусственной эволюции". Само его существование и широкое практическое применение является отличным доказательством работоспособности дарвиновской модели. Однако изготовить высокоэффективный белковый фермент с изначально заданными свойствами методом искусственной эволюции — дело довольно долгое и трудоемкое. Поэтому ученые в последние годы прикладывают большие усилия, чтобы не только выявить, но и понять связи между последовательностью аминокислот и функцией белка. Если эта цель будет достигнута, ферменты с заданными свойствами можно будет проектировать, то есть просто вычислять на компьютере искомую последовательность аминокислот. Это сэкономит огромные средства и позволит осуществить настоящую революцию в химической промышленности. Ведь ферменты — чрезвычайно эффективные катализаторы, способные в тысячи раз ускорить синтез чуть ли не любых химических соединений. Вообще белки — удивительные вещества, заключающие в себе немало парадоксов. Как мы уже знаем, многие аминокислоты, входящие в состав белковой молекулы, могут быть заменены или удалены без заметного изменения свойств белка. С другой стороны, замена всего одной-двух аминокислот в активном центре белковой молекулы может привести к радикальному изменению свойств фермента. В большинстве случаев, конечно, фермент от этого портится, снижает производительность или вовсе становится нефункциональным. В других случаях это может привести к модификации или даже полной смене функции, то есть к появлению нового фермента. Разнообразие белков началось с немногих исходных "мотивов". Некоторые ученые предполагают, что на самых ранних этапах становления жизни — возможно, еще в РНК-мире — методом "слепого поиска" было найдено несколько сотен базовых аминокислотных мотивов, соответствующих основным функциям белков. Изначально эти функции были реализованы на основе очень коротких белковых молекул (пептидов), причем эффективность функционирования была поначалу низкой. Но все равно это был большой шаг вперед по сравнению с неуклюжими рибозимами. Эти первичные "мотивы", оформленные в виде коротких пептидов, в дальнейшем легли в основу всех ныне существующих надсемейств белков. Конечно, разумное просчитанное проектирование может значительно ускорить создание новых белков, на которое эволюции требуются миллионы лет. Иногда, впрочем, белки с новыми функциями возникают в природных популяциях живых организмов гораздо быстрее — буквально за считанные годы или даже месяцы. Здесь нужно различать две принципиально разные ситуации. Иногда новые белки возникают благодаря особым механизмам, специально для этого существующим в клетке. Таковы, например, образование новых антител в ответ на проникновение в организм новой инфекции или целенаправленное изменение поверхностных белков у некоторых патогенных бактерий, имеющее целью обмануть иммунную систему хозяина. Эти особые механизмы направленного изменения генома сами являются результатом длительной эволюции и будут рассмотрены нами отдельно в главе "Управляемые мутации". Но новые белки иногда могут быстро возникать и по обычной, классической схеме — за счет случайных мутаций и отбора. Самый известный пример такого рода — поразительно быстрая выработка бактериями устойчивости к антибиотикам. Как выяснилось, бактерии предпочитают не пассивную, а активную защиту: они не пытаются "обойти" или видоизменить тот биохимический процесс, по которому "бьет" новый антибиотик, а вырабатывают новый фермент, способный активно атаковать и обезвреживать лекарственное средство. Хотя возникновение подобного фермента требует не одной мутации, а нескольких, классическая эволюционная модель вполне способна объяснить это явление. В слаженном появлении целого комплекса необходимых мутаций нет ничего противоречащего простейшей схеме "единичные случайные мутации плюс отбор". Кроме того, на примере развития устойчивости к антибиотикам можно понять, как работает один из механизмов, ограничивающих число допустимых эволюционных траекторий движения к заданной цели. Одним из следствий подобных ограничений могут быть параллелизмы — независимое возникновение сходных признаков в разных эволюционных линиях. Параллелизмом, или параллельной эволюцией, называют независимое появление похожих признаков у разных организмов. Параллелизмы очень широко распространены в природе, и биологи-теоретики уделяют много внимания этому явлению. В нем отчетливо проявляется закономерный (неслучайный) характер эволюции. Одной из основных причин параллелизмов является ограниченность возможных путей эволюционного преобразования организма. Сформулированный Н. И. Вавиловым "Закон гомологических рядов в наследственной изменчивости" (http://evolbiol.ru/vavilov.htm) позволил связать это явление с закономерностями внутривидовой изменчивости. У близких видов встречаются одинаковые вариации — например, у большинства злаков есть остистые и безостые формы. Палеонтология дает много ярких примеров схожести путей эволюции в разных группах (мы поговорим об этом подробно в главе "Направленность эволюции").
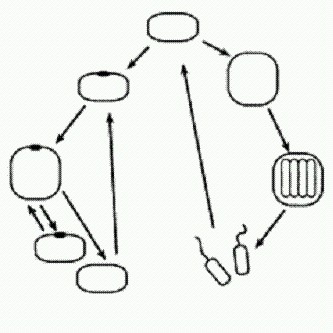
Яркий пример параллельной эволюции: сумчатый саблезубый тигр Thylacosmilus из плиоцена Южной Америки (вверху) и "обычный* саблезубый тигр Smilodon из плейстоцена Северной Америки. Пути эволюции предопределены на молекулярном уровнеВ наши дни бурное развитие молекулярной биологии привело к тому, что многие важные биологические закономерности, в том числе явление параллельной эволюции, временно оказались как бы за рамками "настоящей серьезной науки" — просто потому, что их пока не удается объяснить на молекулярном уровне. Поэтому предпринятая учеными из Гарвардского университета попытка найти молекулярные основы канализированности (ограниченности возможных путей) и повторяемости эволюции имеет большое теоретическое значение (Daniel M. Weinreigh, Nigel F. Delaney, Mark A. DePristo, Daniel L. Hartl Darwinian evolution can follow only few mutational paths to fitter proteins // Science. 2006. v.312. p.111-114). В качестве модели исследователи выбрали адаптацию бактерий к антибиотикам — сравнительно простой эволюционный процесс, высокодетерминированный и повторяемый и к тому же имеющий большое практическое значение. Исследование наглядно показало, что в эволюции может быть реализована лишь очень небольшая часть из общего числа теоретически существующих путей "из точки А в точку Б". Бактерии приспосабливаются к антибиотикам из группы бета-лактамов (к которым относится, в частности, пенициллин) благодаря изменениям гена, кодирующего фермент бета-лактамазу. Предполагается, что мутации возникают случайно, причем вредные мутации отсеиваются отбором, а полезные закрепляются. В ходе адаптации бактерий к цефотаксиму — антибиотику третьего поколения из группы бета-лактамов — в исходный вариант гена бета-лактамазы вносится пять вполне конкретных мутаций, в результате чего устойчивость к антибиотику возрастает в 100 000 раз. Но такой эффект дают только все пять мутаций вместе. Понятно, что одновременное появление сразу пяти "нужных" мутаций невероятно: они должны появляться и фиксироваться последовательно, одна за другой. Значит, на пути к конечной цели организм должен пройти через четыре промежуточных состояния. Если хотя бы одно из них окажется менее выгодным, чем предыдущее, оно будет забраковано отбором, и конечная цель не будет достигнута. Теоретически существует 5(!) = 120 различных траекторий движения от исходного состояния (отсутствие устойчивости к цефотаксиму) к конечному, то есть к очень высокой устойчивости. Экспериментальным путем ученые установили, что из 120 теоретически возможных путей последовательного приобретения пяти мутаций большинство (102) вообще не могут реализоваться, так как требуют на каком-то этапе временного снижения приспособленности (в данном случае под "приспособленностью" понимается устойчивость к цефотаксиму). Оставшиеся 18 путей очень сильно различаются по вероятности своей реализации. Расчеты показали, что в 99% случаев эволюция "выберет" один из 10, а в 50% случаев — один из двух наиболее вероятных путей. Вполне возможно, что это правило распространяется и на эволюцию других белков. Это значит, что молекулярной эволюции свойственны высокие повторяемость и предсказуемость. Разные организмы должны независимо друг от друга двигаться по одним и тем же "разрешенным" эволюционным траекториям. Не исключено, что аналогичные ограничения могут направлять и канализировать дарвиновскую эволюцию и на более высоких уровнях организации живого. Приключения Protozoon (модель возникновения сложного организма из простого)Попробуем понять, как же все-таки в ходе эволюции сложное может рождаться из простого. Для этого я хочу предложить вниманию читателей забавную мысленную модель. Героем нашего повествования будет вымышленное, но довольно правдоподобное одноклеточное существо Protozoon, имеющее вполне обычный жизненный цикл, всего один "орган" — глазок — и всего один внутренний регуляторный фактор (сигнальное вещество X) с двумя эффектами. Это незамысловатое простейшее, как мы увидим, в результате одной- единственной случайной мутации автоматически может дать начало целому вееру сложных многоклеточных форм со своими онтогенезами (способами индивидуального развития), жизненными циклами, разделением тела на вегетативную и генеративную части ("сому" — тело и "герму" — половые клетки). Итак, прошу познакомиться: Protozoon, диплоидный (т.е. имеющий двойной набор хромосом. Таковы все клетки животных, кроме половых. Половые клетки имеют одинарный набор хромосом и называются гаплоидными. У растений и простейших встречаются различные варианты чередования гаплоидных и диплоидных стадий в жизненном цикле. Прокариоты гаплоидны) одноклеточный эукариотический организм с простым жизненным циклом. В благоприятных условиях у зиготы (диплоидной клетки, образовавшейся при слиянии двух гаплоидных половых клеток) образуется глазок. Глазок стимулирует выработку сигнального вещества X.
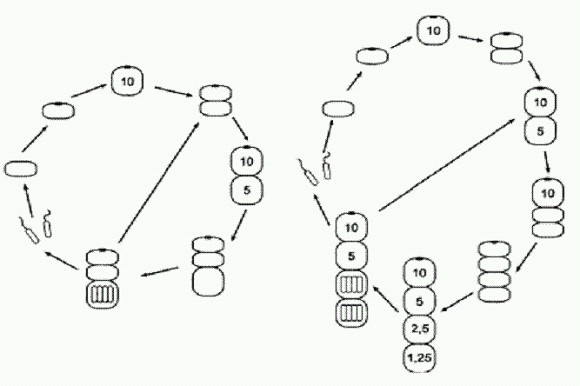
Как у большинства реальных одноклеточных эукариот, клетка нашего Protozoon может делиться двумя способами: митозом (получаются две диплоидные дочерние клетки) и мейозом (получаются четыре гаплоидные половые клетки — гаметы, снабженные жгутиками и потому подвижные). Сигнальное вещество X имеет два эффекта. В концентрации 1 оно подавляет процесс образования глазков, а в концентрации 4 и выше приводит к тому, что созревшая клетка будет делиться митозом (при меньшей концентрации — мейозом). Вещество X способно с определенной скоростью проникать сквозь мембрану (оболочку) клетки в обе стороны. Митоз — деление эукариотической клетки, в результате которого из одной родительской клетки получаются две дочерние с таким же числом хромосом, как и у родительской. Например, митоз диплоидной клетки приводит к формированию двух диплоидных клеток. Мейоз — "редукционное деление" эукариотической клетки, в результате которого число хромосом сокращается вдвое. Из диплоидной родительской клетки (с двойным набором хромосом) получаются четыре гаплоидных клетки с одинарным набором хромосом. V животных таким путем образуются половые клетки — яйцеклетки и сперматозоиды. Глазок к тому же придает клетке полярность. Это проявляется в том, что, когда клетка делится путем митоза, плоскость деления всегда располагается перпендикулярно тому диаметру клетки, на котором находится глазок. В результате глазок "достается" одной из дочерних клеток, а другая поначалу не имеет глазка. Будем считать, что ориентация плоскости деления задается глазком при помощи выделяемого им вещества X. Концентрация этого вещества максимальна возле глазка и постепенно снижается по мере удаления от него. Иными словами, существует "градиент концентрации" вещества X, и плоскость деления всегда перпендикулярна направлению этого градиента. При неблагоприятных условиях зигота не образует глазка, вещество X не вырабатывается, и поэтому, если ей удастся дорасти до зрелости, она делится только мейозом и образует четыре подвижные гаметы. Посмотрим теперь, в чем смысл этого нехитрого регуляторного контура и как складывается жизнь Protozoon в естественных условиях. Если все хорошо и пищи много, все клетки в сообществе Protozoon образуют глазок и делятся митозом. По мере роста плотности популяции увеличивается концентрация X в воде. Это вещество, как мы знаем, проходит через мембрану. Поэтому в конце концов в молодых клетках, еще не успевших образовать глазок, концентрация вещества X (впитанного из окружающей среды) достигнет значения 1. Тогда у них не образуется глазок, и они будут делиться мейозом, гаметы расплывутся, популяция разредится, и произойдет расселение. Очень несложная, вполне правдоподобная и устойчивая система. Но вот у одной особи возникла мутация: утрачена способность клеток расходиться после митоза. Условия благоприятны. Что теперь будет? Сразу, без всякого отбора и дополнительных мутаций, появляется множество разных многоклеточных организмов со своими онтогенезами и жизненными циклами.
Это самый простой из возникших вариантов. Двуклеточный организм, у которого передняя клетка — вегетативная ("сома") с глазком, а задняя — генеративная ("герма"). Задняя клетка, созревая, производит гаметы и сразу же заменяется новой, образовавшейся в результате деления головной клетки. В чем секрет? Оказывается, все дело в сигнальном веществе X. Как только у клетки появляется глазок, концентрация вещества X в ней почти мгновенно достигает максимального уровня 10. Избыточные количества вещества X интенсивно откачиваются из клетки и... проникают в соседнюю, "прилипшую" сзади вторую клетку. И там устанавливается постоянная концентрация X, равная 5. Раньше, когда клетки расходились сразу после деления, подобных ситуаций не возникало. Только теперь, когда клетки остались слипшимися, стало ясно, что проницаемость мембраны для вещества X такова, что в клетке, примыкающей к своей "соседке" с глазком, устанавливается половинная концентрация вещества X. Если к клетке с концентрацией 5 "прилипнет" еще одна клетка без глазка, то в ней установится концентрация 2,5, и так далее, каждый раз вдвое меньше. Раньше это свойство клеточных мембран Protozoon не имело существенного значения, но теперь оно становится ключевым фактором формообразования. Обратите внимание, что в данном случае в созревшем двуклеточном организме (когда и передняя, и задняя клетка уже готовы поделиться) первой всегда делится передняя клетка. Это принципиальный момент. Из-за этого в задней клетке, которая теперь стала третьей по счету, концентрация X быстро снизилась до 2,5 — а при такой концентрации клетка делится уже не митозом, а мейозом. Мы помним, что у исходного одноклеточного существа вещество X регулировало деление клеток: при концентрации выше четырех клетка делилась митозом, при меньшей концентрации — мейозом. Но для одноклеточного существа были безразличны небольшие индивидуальные различия в скорости начала клеточного деления. Между тем концентрация X (Рх) немного влияла на эти скорости. В этом нет ничего удивительного, ведь в конечном счете в клетке все процессы хоть немного, но влияют друг на друга. По этому признаку — характеру влияния X на время начала деления — существовала индивидуальная изменчивость, но пока организм был одноклеточным, эти различия ни на что не влияли, и их было трудно даже заметить. У той особи, из которой в результате мутации сформировался двуклеточный организм, митоз шел быстрее всего при концентрации X, близкой к 10. Поэтому первой стала делиться передняя клетка, а не задняя.
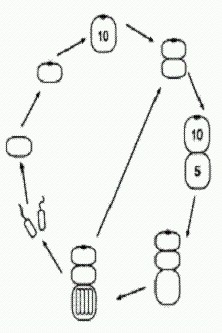
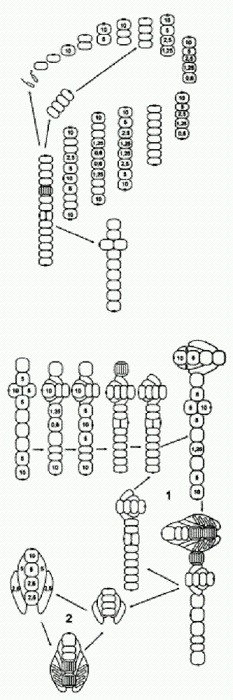
А что будет, если мутация "нерасхождения" произойдет у другой особи, у которой оптимум митоза случайно оказался ближе к Рх = 5? Тогда строение многоклеточного организма окажется еще более сложным. Взрослый организм теперь оказывается четырехклеточным! Он ведет себя как целое и обладает сложным детерминированным (предопределенным) онтогенезом, в ходе которого отдельные части эмбриона определенным образом влияют друг на друга — точно так же, как у настоящих эмбрионов. Такие взаимные воздействия называют "морфогенетическими корреляциями". Более того, онтогенез этого существа обладает еще одним характерным свойством "настоящего" онтогенеза — эквифинальностью, или помехоустойчивостью, то есть способностью компенсировать отдельные нарушения и все равно приходить к той же самой конечной цели. Например, что будет, если приплывет хищная амеба и откусит у молодого четырехклеточного организма головную или хвостовую клетку? Нетрудно рассчитать, что уже через несколько шагов последствия травмы будут полностью компенсированы и жизненный цикл пойдет по исходному сценарию. То же самое справедливо и для стадии двухклеточного организма. Между прочим, недавние исследования показали, что у реальных животных регенерация — восстановление утраченных частей тела — управляется той же молекулярной "программой", которая руководит развитием этих частей в онтогенезе (см. сюжет "Разгадан механизм регенерации конечностей" в главе "Животные"). Кажется, наше выдуманное существо очень понятно объяснило нам, почему так получается и как это работает. Но это еще далеко не все. В созревшем четырехклеточном организме мейоз генеративных клеток произошел раньше, чем митоз второй соматической клетки (у которой Рх = 5). Но у других особей Metazoon (так мы назовем эти новые многоклеточные организмы) в такой ситуации митоз второй соматической клетки может произойти раньше, чем мейоз генеративных клеток. Как и в предыдущем случае, мы предполагаем, что все это было заложено в генофонде исходной популяции Protozoon в виде случайных вариаций, абсолютно несущественных для него (нейтральных), пока он был одноклеточным. Кто же мог знать, что такие пустяки вдруг станут жизненно важными в будущем.
Не забывайте об изменчивости! Как недавно выяснили биологи — преподаватели вузов, одна из основных причин недопонимания эволюционных идей у студентов состоит в том, что многие из них на интуитивном уровне считают всех представителей одного и того же вида одинаковыми. А преподаватели порой забывают в явном виде указать им, что это не так. Я тоже чуть не забыл. Так вот, очень важно всегда помнить, что каждая особь уникальна, все живые существа хоть немножко, но разные. Поэтому отбору всегда есть из чего выбирать. Изменчивость живых организмов, относящихся к одному и тому же виду, связана с тем, что многие гены в популяции существуют в виде нескольких вариантов (их называют аллельными вариантами, или аллелями). Набор генов у каждого представителя данного вида один и тот же, а вот комбинации аллелей у всех разные.
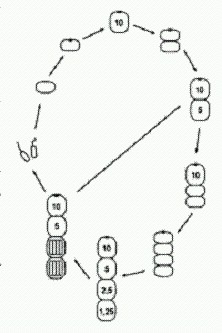
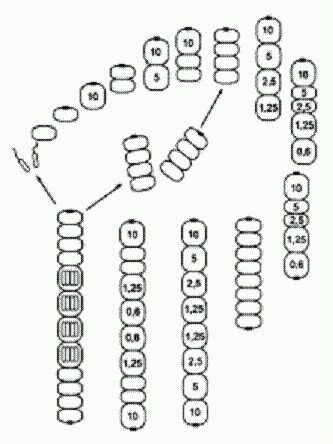
Итак, рассмотрим Metazoon, у которого при Рх = 5 митоз происходит особенно быстро (так, что не успевает произойти мейоз в одновременно созревших клетках). В этом случае неизбежно возникает восьмиклеточный организм, а заодно и новый способ бесполого размножения (на рисунке все идет по часовой стрелке). Фокус тут в том, что, когда клеток в цепочке становится пять, в последней из них Рх снижается настолько, что образуется глазок! И вся картина сразу меняется. Как видим, двухклеточная фаза выпала из основного цикла. Теперь она осталась только в "раннем онтогенезе". После выхода гамет образуются два молодых четырехклеточных организма (бесполое размножение). Но и это еще не все — нашу модель можно достраивать и дальше, производя (мысленно) все более и более сложные многоклеточные организмы на основе простых и вполне реалистичных допущений о характере нейтральной изменчивости у исходного Protozoon. В последнем рассмотренном случае, когда в двух центральных клетках организма (на стадии 10 клеток) Рх упала ниже критического уровня 1, глазок не образовался. Видимо, это произошло потому, что механизм образования глазка не срабатывает, если клетка контактирует более чем с одной соседней клеткой.
Однако вполне можно допустить, что среди исходных Protozoon, опять же в порядке нейтральной и безразличной изменчивости, были и такие особи, у которых механизм образования глазка все-таки мог бы сработать в этой ситуации. Из такого Protozoon, если у него произойдет мутация "нерасхождения", получатся уже не нитчатые (одномерные) а плоские (двумерные) многоклеточные организмы. Здесь глазок появляется только в одной из двух центральных клеток, где Рх упала до о,6, поскольку во второй после этого Рх сразу резко возрастает. Крестообразный десятиклеточный организм становится отныне центральной фигурой, а все предшествующие этапы и циклы — его "ранним онтогенезом". Не перерисовывая эти стадии, проследим дальнейшую, строго детерминированную судьбу нашего десятиклеточного двуглазого Metazoon. Как видим, получился очень интересный онтогенез с двумя замкнутыми циклами. В первом цикле основная жизненная форма — сложный восемнадцатиклеточный организм, который, созрев, производит гаметы, воспроизводит точную уменьшенную копию самого себя и одновременно отпочковывает восьмиклеточный двусторонне-симметричный организм с характерным обликом. Второй цикл состоит в том, что этот восьмиклеточный организм, произведя гаметы, автоматически возвращается к своему исходному строению. Вот такое разнообразие сложных организмов, циклов и онтогенезов образуется само собой из вполне обычного простейшего Protozoon в результате одного небольшого (и очень вероятного) изменения — клетки перестали расходиться после митоза. Ясно, что здесь просчитаны и нарисованы далеко не все возможные варианты. Если допустить, что исходная изменчивость Protozoon по таким не очень важным для него признакам, как пороговые концентрации Рх, вероятность образования глазка в разных непредусмотренных одноклеточным организмом ситуациях, реакция на внешние условия и др., достаточно велика, то число вариантов многоклеточных форм становится очень большим. Однако каждый из вариантов строго детерминирован при данном наборе параметров. Вполне возможно, что в этом разнообразии найдется что "подхватить" отбору. Лишние стадии (например, часть мейозов) могут впоследствии выпасть, заменившись, допустим, апоптозами (смертью клеток); регуляторные механизмы могут усовершенствоваться и даже вовсе замениться; "головные" вегетативные отделы могут постепенно трансформироваться в настоящие головы, взяв на себя особые функции (при этом глазок и индуцируемые им изменения активности генов могут лечь в основу более совершенной системы регуляции онтогенеза). Хочется обратить внимание на интересную особенность восьмиклеточного организма, образующего цикл 2 (на последнем рисунке): пространственное расположение клеток и их плотное "прилипание" друг к другу привели к тому, что клетки дифференцировались на два четких типа, напоминающие настоящие ткани: округлые осевые клетки и сильно вытянутые боковые клетки. Эти последние по своему расположению и форме явно имеют предпосылки в будущем превратиться в покровную ткань (эпителий). Кроме того, в цикле 2 решающую роль играет передача вытянутыми боковыми клетками сигнального вещества X от головной клетки — задним осевым клеткам. Это стабилизирует систему индивидуального развития, выравнивая величину Рх вдоль продольной оси тела и предотвращая образование глазка на заднем конце. Такая функция продольных клеток создает предпосылки для того, чтобы в будущем на этой основе начала развиваться нервная система. В нашей модели (как и в реальном развитии многоклеточных органзмов) каждая отдельная клетка имеет одну и ту же заложенную в нее изначально программу, содержащую несколько условных переходов ("если — то"). Но это вовсе не "программа развития организма", а всего лишь программа поведения отдельной клетки. Совместно развиваясь, клетки меняют входные параметры не только у себя самих, но и у других клеток развивающегося организма, заставляя их тоже менять свое поведение. Именно в этом взаимном влиянии клеток развивающийся организм проявляет себя как целое. Из индивидуальных межклеточных взаимодействий каким-то чудесным образом, без всякого изначального плана и разумного замысла, рождается принципиально новая информация — программа развития организма. Мне эта мысль представляется очень важной для понимания законов развития. И у Metazoon, и в онтогенезе настоящих многоклеточных все происходит настолько закономерно и целенаправленно, что нам кажется, будто развитие идет в соответствии с неким единым, заранее продуманным планом. Но мы видели, что ни у Protozoon, ни у разнообразных произошедших от него Metazoon никакой изначально заложенной программы развития нет и в помине. Эта "программа" рождается сама собой в процессе деления клеток словно бы из ничего. Не так ли обстоит дело и в случае настоящего онтогенеза? А в эволюции? (Идеи такого рода, а также многие другие важные идеи о тесной связи законов онтогенеза и эволюции развивали выдающиеся отечественные эволюционисты, прежде всего И. И. Шмальгаузен, а в наши дни — М. А. Шишкин, разработавший на основе идей Шмальгаузена и Уоддингтона "эпигенетическую теорию эволюции". Разнообразные материалы по этой теории, в том числе труды Шмальгаузена и Шишкина, собраны на страничке http://evolbiol.ru/epigenetics.htm . Там же есть и форум, где все это можно обсудить) Главный вывод, который позволяет нам сделать гипотетический Protozoon и его чудесные превращения, можно сформулировать так: простая живая система может содержать в себе скрытые возможности самопроизвольного усложнения. Опыты, подобные нашему мысленному эксперименту с Protozoon, сейчас активно проводятся и на реальных живых существах. Замечательным объектом такого рода оказалась почвенная бактерия Pseudomonas fluorescens. Появлению многоклеточности мешают обманщики. Экспериментальное изучение эволюции бактерий — одно из перспективных направлений современной микробиологии. Пол Рейни из Оклендского университета (Новая Зеландия) является крупным специалистом в этой области. Один из его любимых объектов — бактерия Pseudomonas fluorescens из группы гамма-протеобактерий. Этот микроб, если предоставить ему необходимый минимум условий, охотно эволюционирует прямо на глазах у исследователей, осваивая новые ниши и вырабатывая различные оригинальные адаптации. В жидкой питательной среде бактерии развиваются сначала как одиночные, подвижные клетки, постепенно занимая всю толщу бульона. Когда в среде становится мало кислорода, получают преимущество бактерии-мутанты, образующие пленку на поверхности среды. Наблюдения за развитием таких колоний навели Рейни на любопытные идеи по поводу происхождения многоклеточности. В многоклеточном организме большинство клеток (так называемые соматические клетки) не передают своих генов следующим поколениям. Они размножаются делением, но только как часть целого организма, который рано или поздно погибает. Соматические клетки — своеобразный "эволюционный тупик", если смотреть на ситуацию с точки зрения индивидуальных клеток. Они жертвуют личными интересами для блага целого — точно так же, как рабочие особи у общественных насекомых.
Pseudomonas fluorescens — подвижная почвенная бактерия, помогающая биологам изучать эволюцию "в пробирке". Вопрос в том, каким образом популяция индивидуальных клеток могла превратиться в единую систему, размножающуюся как целое. Почему естественный отбор перестал эффективно работать на уровне индивидуальных клеток и стал действовать на уровне клеточных популяций? В основе этого превращения, конечно, лежит кооперация между клетками, основанная на том, что в определенных ситуациях индивидууму становится выгодно немного поступиться сиюминутными личными интересами ради коллектива. У Pseudomonas "коллектив" образуется из бактерий-мутантов, которые выделяют повышенное количество веществ, способствующих склеиванию клеток. Такие бактерии после деления не могут "отклеиться" друг от друга. Фокус тут в том, что одиночные клетки барахтаются в толще бульона, а склеившиеся всплывают на поверхность, где кислорода гораздо больше. В результате образуется пленка из бактерий на поверхности среды. Производство клея — дело дорогостоящее, однако общая награда (кислород) с лихвой покрывает расходы.
Колония бактерий-мутантов Pseudomonas fluorescens на поверхности питательной среды. Возникновение подобных колоний — уже большое эволюционное достижение, но до настоящего многоклеточного организма тут еще далеко. Колонии недолговечны, а главное, неспособны размножаться как единое целое. Отбор в них по-прежнему действует на индивидуальном, а не на групповом уровне. Он благоприятствует клеткам-"жуликам", которые перестают производить клей, однако продолжают пользоваться преимуществами жизни в группе. Нет никаких механизмов, которые препятствовали бы такому жульничеству. Безнаказанность способствует быстрому размножению обманщиков, что вскоре приводит к разрушению колонии. Рейни предположил, что ключевым моментом в возникновении многоклеточного организма должна быть выработка механизма борьбы с клетками-обманщиками. Но как может возникнуть такой механизм, если не работает отбор на уровне групп? Чтобы заработал групповой отбор, колония должна научиться размножаться как единое целое — иными словами, должно произойти разделение клеток колонии на вегетативные (сому) и генеративные (герму). Колонии бактерий, о которых идет речь, представляют собой как бы одну сплошную сому, то есть эволюционный тупик. Таким образом, для возникновения многоклеточности должны быть выполнены три условия: 1. кооперация; 2. механизм борьбы с обманщиками; 3. коллективное размножение. Причем все три эволюционные инновации должны развиться более или менее одновременно, что кажется очень маловероятным. Рейни предлагает следующий сценарий выхода из тупика. Если колония, не способная размножаться как целое, подобна соме, то в клетках-обманщиках можно увидеть прообраз гермы. Хотя их размножение губительно для колонии, они в принципе могут взять на себя роль тех "семян", из которых будут развиваться новые колонии. Обманщики действительно могут уплыть из колонии, когда захотят, — ведь они не приклеены к ней в отличие от "честных" бактерий. Если бы обманщики могли давать начало новым колониям, это создало бы предпосылки для группового отбора, который, в свою очередь, смог бы контролировать количество производимых колонией обманщиков. Допустим, в одних колониях мутации, приводящие к возникновению обманщиков, происходят часто, а в других — редко. Если число обманщиков растет слишком быстро, колония будет очень недолговечной, а с ее распадом теряют все свои преимущества и сами обманщики. Поэтому такая колония в конечном счете произведет меньше обманщиков, то есть оставит меньше потомства, чем та, в которой скорость производства обманщиков ниже. Таким образом, отбор, действующий теперь на уровне групп, в принципе может контролировать темп производства обманщиков, и в результате они из эгоистичных паразитов превратятся в неотъемлемую часть целостного организма — генеративные клетки (герму). Чтобы предложенная модель работала, обманщики должны с высокой частотой мутировать обратно в клетки, выделяющие клей. Иначе никаких новых колоний из них не получится. В лабораторных популяциях, с которыми работает Рейни, такая способность у обманщиков имеется. Впрочем, совершенно ясно, что прогрессивное развитие многоклеточных форм не может далеко продвинуться на основе случайной мутационной изменчивости — пусть даже скорость и направленность появления мутаций регулируются отбором (направленность мутирования может состоять в том, что меняются или перестраиваются совершенно определенные гены, а не все подряд, — это явление широко распространено, — однако характер изменений в этих генах все равно остается случайным). Чтобы предложенная схема заработала эффективно, мутационная изменчивость должна смениться модификационной. Это значит, что клетки должны обрести способность становиться "честными" (соматическими) или "обманщиками" (генеративными) за счет регуляции активности генов, не внося наследуемых изменений в геном. Как справедливо отмечает Рейни, такое преобразование вполне по силам эволюции. Однако нужно добавить, что способности бактериальной клетки к модификационной изменчивости крайне ограниченны по сравнению с эукариотической (ядерной) клеткой. Это объясняется прежде всего тем, что у бактерий нет клеточного ядра и наследственный материал постоянно находится под биохимическим ударом цитоплазматических реакций (см. раздел "Сообщество превращается в организм" в главе "Великий симбиоз"). Так что выработать сложные и эффективные системы регуляции активности генов при помощи разнообразных специализированных регуляторных молекул бактериям очень трудно. Возможно, именно по этой причине прокариотам так и не удалось приобрести настоящую многоклеточность, хотя они многократно подходили буквально вплотную к этому. Эукариоты, по имеющимся оценкам, достигали многоклеточного уровня организации как минимум 24 раза независимо в разных эволюционных ветвях. (Источник: Paul В. Rainey. Unity from conflict // Nature. 2007. V. 446. P. 616.) "Несократимая сложность"В последние годы микробиологи получили и другие результаты, наглядно демонстрирующие принципиальную возможность "рождения сложного из простого" на основе элементарных и, по-видимому, случайных наследственных изменений. В ряде экспериментов удалось показать, что одна-единственная случайная мутация может привести к целому комплексу изменений, в том числе к появлению новых полезных свойств и усложнению структуры бактериального сообщества. Ключевым словом здесь является "сообщество". Новая сложность часто рождается в эволюции в результате взаимного приспособления изначально независимых объектов — организмов, клеток или генов. Существующие бок о бок живые системы "притираются" друг к другу так, что, образно говоря, выпуклости одной из них становятся зеркальным отражением впадин другой. И так незаметно, исподволь, из двух самостоятельных объектов рождается нечто принципиально новое — целостная система, одна часть которой теперь уже не может существовать без другой. Креационисты называют это явление "нередуцируемой (или несократимой) сложностью" и считают, что оно доказывает божественное сотворение всего живого. Примеры "несократимой сложности" окружают нас повсюду. Так, люди очень быстро привыкают к достижениям медицины и техники, становятся зависимыми от них. В недалеком будущем креационисты, пожалуй, смогут "обосновать" тезис о том, что человек был сотворен одновременно с мобильными телефонами и антибиотиками — ведь он без них не может существовать! Неизбежным следствием адаптивного (приспособительного) характера эволюции является то, что "несократимая сложность" насквозь пронизывает всю земную биосферу. Организмы постоянно приспосабливаются к среде, в том числе к среде биотической, то есть к другим живым организмам. Именно поэтому подавляющее большинство живых существ, населяющих нашу планету, не может существовать автономно: для выживания им необходимы другие живые существа. Связи между разными видами организмов сильно различаются по степени жесткости и детерминированности. Наиболее жесткие связи характерны для симбионтов и паразитов, которые не могут жить без своих сожителей или хозяев. В мире прокариот, как мы разобрали в предыдущих главах, чрезвычайно широко распространены симбиоз и связанные с ним явления, такие как альтруизм, самопожертвование и сложное коллективное поведение. Хотя генетика микроорганизмов очень активно изучается (полностью прочтены геномы многих сотен микробов), генетические механизмы становления подобных взаимосвязей остаются во многом загадочными. Интересное исследование провели недавно ученые из Института биологии развития им. Макса Планка (Тюбинген, Германия), которым удалось наглядно продемонстрировать, что одна-единственная мутация может породить способность к сложному коллективному поведению и радикально изменить взаимоотношения между организмами в сообществе (Francesca Fiegna, Yuen-Tsu N. Yu, Supriya V. Kadam, Gregory J. Velicer Evolution of an obligate social cheater to a superior cooperator // Nature. 2006. v.441. p.310-314). Фактически в этом эксперименте было показано, как случайная мутация создает новую "несократимую сложность" без всякого заранее намеченного плана или "разумного замысла". Работа проводилась на почвенной бактерии Myxococcus xanthus, относящейся к группе миксобактерий. Для этих прокариотических организмов характерно сложное коллективное поведение. Например, они иногда собираются в большие скопления и устраивают коллективную "охоту" на других микробов. "Охотники" выделяют токсины, убивающие "добычу", а затем всасывают органические вещества, высвободившиеся при распаде погибших клеток. Как и другие миксобактерии, Myxococcus при недостатке пищи образует плодовые тела, в которых часть бактерий превращается в споры. В виде спор микробы могут пережить голодные времена. Плодовое тело "собирается" из огромного множества индивидуальных бактериальных клеток. Создание такой крупной и сложной многоклеточной структуры требует слаженных действий миллионов отдельных бактерий, из которых лишь малая часть получает прямую выгоду, а все остальные жертвуют собой ради общего блага. Дело в том, что лишь очень немногие из участников коллективного действа смогут превратиться в споры и передать свои гены следующим поколениям. Все остальные выступают в роли "стройматериала", обреченного умереть, не оставив потомства. Человек — животное социальное, и поэтому альтруизм всегда вызывает у нас чувство бурного одобрения (по крайней мере на словах). Но он, однако, имеет и оборотную сторону. Там, где много альтруистов, обязательно заводятся какие-нибудь жулики, обманщики и паразиты, которые рады поживиться за чужой счет, хотя сами ничем для других жертвовать не собираются. Среди миксококков такие обманщики тоже имеются: это генетические линии (или штаммы), не способные к образованию собственных плодовых тел, но умеющие пристраиваться к чужим плодовым телам и образовывать там свои споры. Один из таких штаммов (исследователи назвали его ОС, от obligate cheater — "обязательный обманщик") не способен образовывать споры, когда живет один, однако он успешно внедряется в чужие плодовые тела и спор продуцирует даже больше, чем "альтруистический" штамм-хозяин, построивший плодовое тело. Ученые экспериментировали с этой системой "паразит-хозяин", выращивая смешанную культуру альтруистов и обманщиков попеременно то в "голодной", то в богатой питательными веществами среде и следя за колебаниями численности двух штаммов. Во время голодовок выжить могли только те бактерии, которым удалось превратиться в споры. В одном из опытов в штамме ОС произошла мутация, в результате которой бактерии не только восстановили утраченную способность к самостоятельному (без помощи другого штамма) образованию плодовых тел и спор, но и получили дополнительное преимущество. По не вполне ясным причинам мутантные бактерии оказались защищены от нахлебников — представителей штамма ОС, своих прямых предков. Возникший в результате мутации новый штамм получил условное название РХ (от phoenix — "феникс"). Этот штамм появился и быстро размножился как раз в тот момент, когда почти все бактерии в экспериментальной культуре (исходный "дикий" штамм и паразиты ОС) погибли. Деградация смешанной культуры была вызвана тем, что с каждым экспериментальным циклом доля паразитов неуклонно росла, и в конце концов альтруистов — представителей исходного дикого штамма — осталось слишком мало, чтобы обеспечить себя и других плодовыми телами. Штамм РХ, как выяснилось, побеждает в конкурентном соревновании и диких миксококков, и паразитов ОС. Он не нуждается для образования плодовых тел в других штаммах и защищен от паразитизма. Генетический анализ исследованных штаммов показал, что РХ отличается от своего прямого предка ОС единственной мутацией — заменой нуклеотида в некодирующей (вероятно, регуляторной) области одного из генов. Белок, кодируемый эти геном, судя по всему, влияет на активность каких-то транскрипционных факторов — белков, регулирующих активность генов. Действительно, активность целого ряда генов у РХ сильно изменилась по сравнению с ОС. Паразитический штамм ОС произошел от исходного дикого штамма в результате 14 мутаций, причем все они сохранились у РХ. Таким образом, удивительная мутация, резко повысившая конкурентоспособность бактерий штамма РХ, вовсе не была возвратом к исходному, предковому состоянию: это было настоящее эволюционное новообразование. Данный пример еще раз подтверждает идею, которая в последнее время стала уже практически общепризнанной: небольшие изменения регуляторных генов могут приводить к крупным эволюционным преобразованиям, в том числе к появлению новых свойств и функций и к общему усложнению живой системы. Регуляторный компромиссЭта идея до сих пор была опубликована только в электронном виде на моем сайте "Проблемы эволюции", а такая публикация считается неофициальной, как бы ненастоящей. К моему удивлению, на нее стали ссылаться в капитальных трудах по теории эволюции (С. Н. Гринченко. Системная память живого. 2004.; Ю. В. Чайковский. Наука о развитии жизни. 2006.), из чего я сделал вывод, что пора изложить ее наконец и на бумаге. Идею можно рассматривать как развитие представлений об "адаптивном компромиссе" (А. П. Расницин. Процесс эволюции и методология систематики. 2002. // Труды Русского энтомологического общества. Т. 73. С. 1-108. http://www. palaeoentomolog.ru/Publ/Rasn/methodology.html). Гипотеза "адаптивного компромисса" предложена для объяснения того очевидного обстоятельства, что многие живые организмы весьма далеки от совершенства, их приспособленность относительна, а многим чертам строения вообще не удается приписать никакого адаптивного смысла. Предполагается, что, поскольку жизнь предъявляет к каждому организму множество разнообразных и часто противоречивых требований, задачей эволюции становится поиск оптимального компромисса. Нельзя достичь совершенства во всем сразу, поэтому чем-то приходится жертвовать. Кроме того (и, возможно, это главное), разные части организма, предназначенные для выполнения разных функций, должны соответствовать не только требованиям внешней среды, но и друг другу. Они должны быть взаимно приспособлены, чтобы организм исправно функционировал как целое. Это резко ограничивает спектр возможных (разрешенных) эволюционных преобразований и делает эволюцию отчасти направленной и предсказуемой. Рассмотренная нами модель эволюции вымышленного существа Protozoon позволяет дополнить представления о компромиссе, сделав акцент на проблеме внешних и внутренних регуляторов. Появление необычайного многообразия форм Metazoon стало возможно только благодаря тому, что у исходного Protozoon уже имелась элементарная система регуляции жизненного цикла, включающая один внутренний регуляторный фактор. Под словом "внутренний" я подразумеваю то обстоятельство, что клетка Protozoon, решая, какой из двух вариантов деления ей выбрать (митоз или мейоз), ориентировалась не на внешние факторы (температуру и т.п.), а на внутренние. Она принимала решение в зависимости от концентрации в цитоплазме вещества X, выделяемого глазком. Без этого элементарного внутреннего регулятора никакого подобия целостного организма в нашей модели не получилось бы. По-видимому, наличие внутренних регуляторных связей является необходимым условием прогрессивного развития системы. Например, у нашего Protozoon в хороших условиях целесообразно было выполнение клеткой двух функций: 1. образование глазка и 2. размножение путем митоза. Но если бы каждая из этих функций регулировалась внешними условиями независимо одна от другой, Protozoon не смог бы так легко превратиться в сложный многоклеточный организм. Мы знаем, что у Protozoon внешние условия определяли только развитие глазка, а уже глазок, в свою очередь, вызывал митоз. Таким образом, митоз стал реакцией на состояние не внешней, а внутренней среды организма. Казалось бы, это неудобно. Вдруг после образования глазка условия ухудшатся, и тогда митоз будет нецелесообразен, но все равно произойдет — ведь глазок уже успел появиться. На самом деле хоть это и неудобно, но в значительной мере неизбежно, потому что клетка — это целостная система. Появление глазка меняет структуру и внутреннюю среду клетки. Если в течение долгого времени большинство клеток, совершающих митоз, имели при этом глазок, то в процессе постепенного, медленного совершенствования механизма регуляции митоза у Protozoon должна была произойти "подгонка" этого механизма к ситуации наличия глазка, т. е. именно к такой внутренней среде и структуре клетки, которая определяется глазком. То же самое происходило и с механизмом мейоза. Это своего рода специализация, неизбежно происходящая в стабильных условиях под действием отбора ("стабильность" в данном случае состояла в стабильном, повторяющемся совпадении наличия глазка с митозом, отсутствия глазка — с мейозом). Точно так же, например, внутриклеточные симбиотические бактерии довольно быстро "привыкают" к удобной и обеспеченной жизни внутри клеток хозяина и утрачивают способность жить самостоятельно (см. главу "Великий симбиоз"). Точно так же и мы очень быстро привыкаем к разным новым удобствам и удовольствиям, и чем сильнее привычка, тем труднее потом отвыкать. В результате спустя какое-то время оказалось, что митоз у Protozoon наилучшим образом происходит в клетках с глазком, а мейоз — в клетках без глазка. Гаметы, образующиеся в клетках с глазком, оказываются менее жизнеспособными; то же относится и к молодым клеткам, образовавшимся в результате митотического деления клетки без глазка. Теперь уже очевидной становится целесообразность прочного закрепления связей "глазок-митоз" и "отсутствие глазка — мейоз". Еще очевиднее это у Metazoon. Допустим, отбор благоприятствовал крупным организмам и был закреплен в качестве нормы двойной цикл, показанный на последнем рисунке. Что произойдет, если вдруг восстановится древняя, исходная форма регуляции митоза/мейоза — напрямую внешними условиями? В "плохих" условиях все зиготы будут сразу "разваливаться" на гаметы, и это даст хороший шанс всей популяции погибнуть, т. к. гаметы — самая уязвимая стадия; если же популяция и выживет, то вернется в исходное одноклеточное состояние до тех пор, пока условия не улучшатся. В "хороших" условиях дело пойдет гораздо хуже. Будут расти бесформенные многоглазчатые комки клеток или бесконечные нити, ветвящиеся или нет в зависимости от возможности образования глазков в их середине. Онтогенез и организм как целостные системы будут полностью разрушены. Таким образом, в процессе эволюции должен достигаться своего рода регуляторный компромисс между требованиями адаптивности (соответствия внешним условиям) и требованиями целостности системы. Первая группа требований, диктуемая отношениями организма с внешней средой, стремится увеличить роль внешних регуляций (чтобы адекватно реагировать на изменение условий). Вторая группа требований, диктуемая целостностью онтогенеза и организма, стремится увеличить роль внутренних регуляций (чтобы отдельные части и функции сложной системы, подогнанные друг к другу, развивались и действовали согласованно). У более сложных организмов регуляторный компромисс должен все больше склоняться в сторону преобладания внутренних регуляций (чем сложнее система, тем более опасным становится рассогласование ее частей). Это необходимо чем-то компенсировать, поскольку такое нарастающее "замыкание" регуляций внутри организма ведет к росту несоответствия между требованиями меняющейся среды и поведением живого существа, которое становится саморегулирующимся и самодовлеющим. Как можно это компенсировать? По-видимому, несколькими способами: 1. формированием принципиально новых внешних регуляторных связей, причем новые регуляторы должны действовать не непосредственно на базовые процессы в организме, а на системы их внутренней регуляции; 2. повышением независимости организма от внешних условий путем поддержания внутреннего гомеостаза (температуры тела, рН, осмотического давления и др.), чтобы внешние факторы реже вступали в противоречие с внутренними процессами; 3. искусственным созданием или нахождением для себя подходящих условий (термитники, гнезда, другие укрытия или жилища); преобразованием среды; активным перемещением в места, где условия благоприятны (сезонные миграции птиц, суточные миграции планктона и др). Очевидно, что любой из перечисленных путей требует дальнейшего усложнения организма и онтогенеза. Первый путь вводит новые регуляторные связи — это, так сказать, усложнение "по определению". Второй путь требует прогрессивного развития обмена веществ, покровных тканей — здесь тоже без усложнения всей системы не обойтись. Третий путь требует развития нервной системы — регуляторики самого высокого уровня. В этом можно заметить механизм положительной обратной связи: усложнение системы ведет к конфликту, снятие которого возможно только путем дальнейшего усложнения (Положительные обратные связи (цепные реакции) - почти обязательное условие всех крупных преобразований, происходящих в природе. Это, на мой взгляд, одно из важнейших эмпирических обобщений, которое можно сделать на основе изучения путей и механизмов эволюции. Данное правило очень помогает в работе: если мы видим, что в природе произошло что-то значительное, и хотим понять механизм, то прежде всего следует искать положительную обратную связь). Сняв путем усложнения старые конфликты, организм неизбежно сталкивается с новыми. Система стала сложнее, и, следовательно, поддержание ее целостности требует совершенствования внутренних регуляций, а это, как мы знаем, ведет к новому разрыву между "замыкающейся на себя" живой системой и изменчивой средой. Может быть, в этом состоит одна из причин наблюдаемого ускорения прогрессивной эволюции. Биологи давно заметили эту общую тенденцию: чем сложнее организм, тем быстрее он эволюционирует по пути дальнейшего усложнения. Причины этого до сих пор не вполне ясны. Возникновение и наследование модификаций на примере MetazoonЕще один важный теоретический вопрос, который можно рассмотреть на нашем примере с Metazoon, — это вопрос о возникновении адаптивных модификаций. Так называют способность организма более или менее осмысленно (то есть целесообразно) менять свое поведение, строение и онтогенез в зависимости от внешних условий, причем без изменений генома. Простейший пример — мышцы увеличиваются от упражнения. Модификации — это типичные "благоприобретенные признаки". Синтетическая теория эволюции отрицает возможность наследования модификаций, а ламаркизм основан на признании возможности их наследования. Сам Дарвин в этом отношении был ламаркистом: он считал, что наследование модификаций, возникающих в ходе упражнения органов, вполне возможно. Вопрос о наследовании приобретенных признаков мы рассмотрим в главе 8, а сейчас займемся другой проблемой, может быть, даже более важной: откуда берется сама адаптивная модификация? Ведь ясно, что способность адекватно изменяться в зависимости от условий — это особый сложный механизм, он не может просто "быть заложен изначально" в свойствах живой материи. Способность к полезным модификациям вырабатывается в ходе эволюции, как и любые другие приспособительные свойства организма. В дальнейшем "модифицированное" состояние может зафиксироваться и стать наследственной нормой. Но самое главное — понять, как возникают новые модификации, ведь именно в этот момент происходит эволюционное новообразование. Многие биологи-теоретики справедливо полагают, что превращение модификации в стойкий наследуемый признак — это акт упрощения, а не усложнения. Действительно, организм, способный к модификации, имеет два или более возможных путей развития, из которых он выбирает один в зависимости от условий. Организм, у которого один из этих путей наследственно закрепился и стал единственно возможным, явно проще исходного. Посмотрим, каким образом у Metazoon модификация может развиться на основе случайной изменчивости. Допустим, существует Metazoon, у которого из всего разнообразия потенциально возможных жизненных циклов реализуется только два — "двухклеточный" и "четырехклеточный".
Будем считать, что у этого организма клетки с Рх < 4 приступают к мейозу раньше, чем одновременно созревшие клетки с Рх > 4 приступают к митозу. Допустим, что это свойство уже зафиксировалось в геноме и не подвержено изменчивости. Это позволит четырехклеточному организму не превращаться чуть что в восьмиклеточный, а нам упростит изложение. Какой цикл из двух возможных будет избран, изначально определяется чистой случайностью. Все зависит от того, какая из клеток двухклеточного организма поделится первой — передняя или задняя, а это вполне может быть делом случая. Такую ситуацию легко себе представить. Обе клетки находятся на одной стадии зрелости, поэтому то, какая из них начнет делиться чуть раньше, может зависеть от массы случайных факторов. Теперь представим себе, что в популяции начал действовать отбор. Например, некое хищное существо стало избирательно заглатывать четырехклеточники, не трогая при этом ни двух-, ни одноклеточные стадии. Следовательно, станут избирательно выживать и размножаться те Metazoon, у которых на двухклеточной стадии передняя клетка делится первой и которые поэтому не достигают четырехклеточной стадии. В этой изменившейся ситуации ни нас, ни Metazoon уже не удовлетворят разговоры о "чистой случайности"! Придется разбираться: от чего же все-таки зависит, какая из двух клеток поделится первой? В любой живой клетке все процессы связаны в единую сложную сеть. В конечном счете каждое вещество, каждое химическое превращение и каждый регуляторный эффект через более или менее длинную цепочку промежуточных звеньев влияет на все остальные. Большинство этих влияний — слабые, побочные, не имеющие реального значения. Но мы должны помнить, что на любую функцию клетки — в том числе и на скорость митоза — в той или иной степени влияют тысячи белков (каждый из которых может существовать в популяции в виде нескольких разных вариантов — аллелей) и тысячи "малых молекул", поступающих в клетку извне или образующихся в ней, являющихся субстратами, продуктами и промежуточными этапами обмена веществ. Материалом для отбора являются в первую очередь аллельные варианты генов, кодирующих белки. Жесткий отбор немедленно даст преимущество такому сочетанию этих вариантов, при которых через любую по длине и качеству цепочку взаимодействий сигнал, пусть самый слабый, будет передан из точки А в точку Б регуляторной сети. Здесь "точка А" — это любая из тех особенностей, которые отличают переднюю клетку от задней в двухклеточном организме. "Точка Б" — это сдвиг момента начала митоза (на "чуть раньше" для передней клетки или "чуть позже" для задней). Учитывая огромную сложность регуляторной сети в любой клетке, практически невозможно предсказать, за какой из множества возможных путей передачи сигнала из А в Б зацепится отбор. Это должна быть такая цепочка, реализация (усиление) которой не принесет заметного ущерба жизнеспособности организма. Цепочка может оказаться как короткой, так и длинной. Она может оказаться неожиданной в том смысле, что будут задействованы белки, не имеющие прямого отношения ни к митозу, ни к отличиям передней клетки от задней. Но именно через них пройдет путь передачи сигнала, потому что были выполнены два условия: 1. именно эта цепочка первой "подвернулась под руку" естественному отбору. Или, говоря строже, среди имеющихся в популяции генетических вариантов оказались такие, в которых именно по этой цепочке из точку А в точку Б уже передавался слабенький "сигнал", в результате чего митоз передней клетки двуклеточного организма чуть ускорялся (или чуть замедлялся митоз задней клетки); 2. "усиление" этой регуляторной цепочки под действием отбора не вызвало вредных побочных эффектов. В нашем случае известно, что передняя клетка отличается от задней вдвое большей концентрацией вещества X и что уже существует регуляторная система, определяющая выбор способа деления в зависимости от концентрации X (следовательно, есть "датчики", регистрирующие эту концентрацию). С большой вероятностью эти готовые блоки будут задействованы в формировании новой регуляторной связи. Эволюция создает новые структуры из того, что под рукой. Поначалу некоторым регуляторным белкам, возможно, придется взять на себя новую функцию вдобавок к старой; в дальнейшем гены этих белков могут дуплицироваться (удвоиться), и произойдет разделение функций. Очень важно понять, что для формирования принципиально нового (чуть ли не любого!) регуляторного эффекта в организме, скорее всего, уже есть преадаптации — своего рода "заготовки". Это следует из самого факта взаимосвязанности всех клеточных процессов. Но мы пока еще не пришли к настоящей адаптивной модификации. Мы рассмотрели только процесс наследственного закрепления одного из двух "случайных" вариантов онтогенеза. Такой процесс соответствует представлениям о стабилизирующем отборе (И. И. Шмальгаузен. Факторы эволюции. Теория стабилизирующего отбора. 1968. http://evolbiol.ru/factory.htm.) (произошла "стабилизация" онтогенеза) и ведет к снижению изменчивости. У нас был организм с двумя возможными вариантами развития, один из которых выбирался случайным образом. Теперь мы пришли к организму, у которого всегда выбирается один и тот же вариант (двухклеточный). Этот организм отличается от исходного тем, что у него возникла новая регуляторная связь, новый "оператор условного перехода" в программе поведения клетки. Примерно такой: "если я — передняя из двух клеток, то я делюсь быстрее". Стал ли организм сложнее исходного, возникла ли новая сложность? На мой взгляд, да. Мне представляется, что именно формирование новой регуляторной связи в программе поведения клетки и является базовым элементарным актом усложнения организма. Однако если смотреть не на программу, а на результат ее выполнения, то есть на сам организм, то усложнение в данном случае выглядит неочевидным. Можно даже подумать, что возможности организма сузились, ведь из двух имевшихся вариантов развития остался только один. Чтобы усложнение стало совсем уж бесспорным, мы должны сделать еще один шаг и посмотреть, каким образом организм, родившийся в нашем предыдущем мысленном эксперименте, вырабатывает способность к настоящей адаптивной модификации. Оказывается, для этого достаточно создать еще одну регуляторную связь. Допустим, тот хищник, который избирательно поедает четырехклеточные организмы, активен только при определенных условиях среды — например, при затяжных дождях (назовем эти условия У1). При иных условиях — например, при ясной погоде (У2) — хищник не опасен для Metazoon. Допустим, он откочевывает в другую часть водоема, где Metazoon не живет, и там охотится на кого-то другого. И в условиях У2 преимущество получают четырехклеточные, а не двухклеточные особи Metazoon. В такой ситуации создаются предпосылки для возникновения новой регуляторной связи, которая будет передавать сигнал от какого-нибудь "датчика внешних условий" к той системе регуляции скорости митоза в зависимости от положения клетки в двухклеточном организме, которая возникла до этого. "Входными данными" для этого нового регуляторного контура будут служить внешние условия (У1 или У2). Скорее всего, эти условия хоть как-нибудь, хоть немного, но будут отражаться на внутренней среде клетки, поэтому отбору будет за что зацепиться. В качестве одного из вариантов можно предположить, что сигналом о наступлении условий У2 будет служить отсутствие запаха хищника. На выходе новый регуляторный контур должен взаимодействовать с одним из элементов старого контура. В случае наступления условий У2 новый контур будет просто-напросто блокировать старый, отключать его. Программа поведения клетки будет теперь выглядеть так. Если условия = У1, то если я — передняя из двух клеток, то я делюсь быстрее. Таким образом, теперь система ускорения митоза в передней клетке будет срабатывать только в том случае, если условия = У1 (или, что то же самое, если условия не равно У2), то есть в случае присутствия хищника. Если же хищника нет, ускорение митоза в передней клетке блокируется и восстанавливается исходная ситуация случайного выбора одного из двух вариантов развития. Это уже самая настоящая способность к адаптивной модификации, хотя и не доведенная до полного совершенства. Если хищник присутствует, все особи развиваются по двухклеточному пути; если хищника нет, то часть особей развивается по двух-, а часть — по четырехклеточному пути. Думаю, что читатели, особенно те из них, кто немного знаком с программированием, легко смогут сами придумать, как довести эту систему до совершенства, то есть заставить организмы в условиях У1 (когда хищника нет) всегда выбирать четырехклеточный путь развития. Похоже на то, что, добавляя понемножку новые "если" в программу поведения клетки, можно получить из нашего Metazoon организм чуть ли не любой степени сложности. Мне кажется, на этой основе можно было бы сделать занятную компьютерную игру. Теперь уже никак нельзя сомневаться в том, что на наших глазах система стала сложнее — произошло рождение новой сложности. По-видимому, именно таким способом — путем создания дополнительных регуляторных контуров, новых "операторов условного перехода" в программе поведения клетки — усложнялось в ходе эволюции строение многоклеточных организмов, в особенности животных, о которых пойдет речь в следующей главе. Перейти от достигнутого нашим Metazoon высокоразвитого состояния к одному фиксированному жизненному циклу очень просто. Для этого не надо создавать ничего нового, напротив, нужно упростить систему. Это называют "генокопированием модификаций". В результате "генокопирования" один из вариантов развития будет жестко закреплен в геноме. Он начнет реализовываться всегда, вне зависимости от условий. Представим себе, что условия У2 (хищника нет) перестали возникать, среда стала более стабильной: всегда существуют только условия У1 (хищник есть). Четырехклеточный вариант жизненного цикла становится ненужным. Регуляторный контур, обеспечивающий отключение ускорения митоза передней клетки в отсутствие хищника, больше не задействуется и просто отмирает под грузом случайных мутаций. Мы вернулись к тому же состоянию, которое существовало после первого этапа нашего мысленного эксперимента (когда первая регуляторная связь уже сформировалась, а вторая — еще нет). Регуляторную цепочку можно представить себе как последовательность переключателей, где изменение регистра первого переключателя приводит к переключению следующего, и т. д. Если в течение многих поколений все переключатели остаются в одном и том же положении, они "ржавеют" и застопориваются в нем навсегда, а затем могут и вовсе "рассосаться". Случайные мутации и отбор — точнее, в данном случае, его отсутствие, поскольку особи с "ржавыми" переключателями перестают выбраковываться — прекрасно объясняют этот процесс упрощения. В конце концов остается последний "ржавый", навсегда включенный переключатель, вплотную примыкающий к регулируемому процессу. В нашем случае это будет выражаться в том, что в определенном месте регуляторной цепочки, идущей от "порядкового номера клетки" к митозу — а именно там, где раньше эта цепочка сливалась с другой, идущей от "внешних условий", — будет стоять "ржавый переключатель", якобы показывающий, что условия = У1 Вот у нас и унаследовалась модификация. Правда, не сразу ("по Ламарку"), а за ряд поколений ("по Дарвину"). Как видим, ничего хитрого в механизме наследования модификаций нет, одно сплошное упрощение.
Почему эволюция иногда сохраняет "ненужные" регуляторные контуры? Любопытно, что в некоторых случаях регуляторные контуры могут в течение миллионов лет сохранять способность адекватно реагировать на изменения положения тех "переключателей", которые давным-давно "заржавели" или вовсе исчезли. Современные методы иногда позволяют "воскресить" отмерший переключатель, и тогда появляется возможность наглядно убедиться в том, что система по-прежнему в рабочем состоянии. Таким способом, например, удалось вырастить зачатки зубов у куриного эмбриона, воздействовав на ткани челюсти соответствующими регуляторными веществами, взятыми у других животных. Предки курицы утратили зубы десятки миллионов лет назад, но многие компоненты регуляторного контура, необходимого для роста зубов, сохранились! Почему они не рассыпались под грузом случайных мутаций? Дело в том, что у животных важнейшие блоки регуляторных каскадов являются многофункциональными, то есть входят в разных комбинациях в состав множества регуляторных контуров. Поэтому отбор оберегает эти блоки, отсеивая мутации, ведущие к их повреждению. Еще один пример "воскрешения отмершего переключателя" мы рассмотрим в главе "Животные" (см. сюжет "Разгадан механизм регенерации конечностей"): активировав один-единственный регуляторный ген, ученым удалось вернуть куриному эмбриону способность к регенерации конечности, давным-давно утраченную предками птиц. Что же касается нашего Metazoon со способностью к адаптивной модификации, то он — существо весьма перспективное. Возникшие у него новые регуляторные каскады открывают великолепные возможности для дальнейшей эволюции. Сначала новшества могут появляться за счет мелких мутаций в регуляторных генах, чуть меняющих реакцию белков на различные сигналы, или в результате попадания организма в непредвиденную ситуацию, на которую не были рассчитаны регуляторные связи и в которых они начнут работать немного иначе. В дальнейшем эти новые "случайные отклонения" могут закрепляться или как основная норма, или как новая адаптивная модификация.
Выведена гусеница, меняющая цвет при нагревании. Недавно возможность появления новой адаптивной модификации (полифенизма) под действием отбора была продемонстрирована экспериментально. Американские ученые путем искусственного отбора вывели гусениц, цвет которых дискретно меняется в зависимости от температуры: в тепле развиваются зеленые гусеницы, на холоде — черные. Норма реакции и полифенизм. Многие далекие от генетики люди полагают, что в генах "записано" строение организма (генотип определяет фенотип). Это не совсем так. В действительности генотип определяет не фенотип как таковой, а норму реакции — определенный спектр возможностей развития. Какая из этих возможностей будет реализована, зависит уже не от генов, а от условий, в которых будет происходить развитие организма. Обычно эти предусмотренные генотипом допустимые варианты фенотипа образуют непрерывный, плавный ряд, но иногда они бывают дискретны. В таком случае говорят о явлении полифенизма. Это значит, что при одном и том же генотипе в зависимости от условий будет реализован один из нескольких дискретных вариантов фенотипа. Например, из муравьиного яйца с одним и тем же генотипом могут развиться и рабочий муравей, и крылатая самка — в зависимости оттого, как будут кормить личинку. Другие примеры полифенизма — одиночная и стайная формы перелетной саранчи, бескрылая и крылатая формы тлей. Полифенизм — одна из форм адаптации к переменчивым условиям среды. Известно, что пороговые значения внешних факторов, вызывающие "переключение" развития на альтернативный путь, могут меняться под воздействием отбора. Однако сам механизм этого переключения пока мало изучен.
В качестве объекта выбрали бабочку Manduca sexta. У этого вида полифенизма нет, однако он есть у близкородственного вида М. quinquemaculata, у которого при 20 °С развиваются черные гусеницы, а при повышении температуры до 28 °С — зеленые.
Цвет гусениц после теплового шока в ходе отбора становился все более зеленым в полифенной линии и все более черным — в монофенной. У Manduca sexta вне зависимости от температуры гусеницы зеленые, однако встречаются и мутанты — черные гусеницы. Но это не полифенизм, а обыкновенная наследственная изменчивость (полиморфизм), поскольку цвет гусеницы определяется генотипом, а не внешними условиями. Мутация black ("черная окраска") приводит к уменьшению выработки так называемого ювенильного гормона, что в свою очередь ведет к меланизации (почернению) покровов. Нормальные (не мутантные) зеленые гусеницы не меняют свой цвет ни при каких обстоятельствах, однако ученые обнаружили, что окраска черных мутантов менее стабильна. Если подвергнуть гусеницу- мутанта в определенном возрасте шестичасовому тепловому шоку (нагреванию до 30-33°С), то после очередной линьки гусеницы получаются разные — черные, с прозеленью или почти нормального зеленого цвета. Ухватившись за это обстоятельство, ученые приступили к искусственному отбору гусениц с мутацией black. В одной линии отбирались гусеницы, реагирующие на тепловой шок минимально (т. е. остающиеся черными или почти черными). В другой линии, наоборот, отбирались те особи, которые реагировали на перегрев максимальным позеленением. Уже в седьмом поколении в первой линии была полностью утрачена способность реагировать на тепловой шок изменением окраски (полный монофенизм). Во второй линии за 13 поколений возник настоящий полифенизм. Все гусеницы в этой линии теперь становились ярко-зелеными при постоянной температуре 28°С. Теплового шока, нагревания до 30-33°С, уже не требовалось. При более низких температурах гусеницы оставались черными. Чтобы проверить, связано ли изменение окраски у полифенных гусениц с ювенильным гормоном, ученые провели следующий жестокий эксперимент: туго перевязали гусениц поперек брюшка, чтобы гормон, синтезирующийся в голове и груди, не мог попасть в заднюю часть тела. Гусеницы стоически перенесли издевательство и доказали правильность предположений своих мучителей, позеленев спереди и оставшись черными сзади. Гусеницы из монофенной линии, естественно, оставались черными хоть с перевязками, хоть без. Результаты продемонстрировали тот путь, который проделывает случайная мутация (такая как мутация black) под действием отбора до настоящего полифенизма, и что изменения гормональной регуляции могут играть важную роль в подобных эволюционных преобразованиях. Более общий эволюционный вывод, который можно сделать на основании этой работы, состоит в том, что генотип может содержать в себе в скрытом виде разнообразные новые возможности (варианты развития), до поры до времени не проявляющиеся, замаскированные различными регуляторными механизмами — например, той же гормональной регуляцией. Но в экстраординарных условиях, в результате мутации или стресса, регуляторные механизмы могут дать сбой, и тогда скрытая изменчивость станет явной и может стать основой для развития новых полезных адаптаций. (Источник: Suzuki Y., Nijhout Н. F. Evolution of a Polyphenism by Genetic Accommodation // Science. 2006. V. 311. P. 650-652.) Что почитать на эту тему в ИнтернетеН. А. КОЛЧАНОВ, В. В. СУСЛОВ. Кодирование и эволюция сложности биологической организации. 2006. // "Эволюция биосферы и биоразнообразия" (к 70-летию А. Ю. Розанова), http://evolbiol.ru/r_kolchanov.htm Н. А. КОЛЧАНОВ, В. В. СУСЛОВ, В. К. ШУМНЫЙ. Молекулярная эволюция генетических систем. 2003. http://ev0lbi0l.ru/kolchanov2003.htm А. В. МАРКОВ. 2003-2007. Обзоры и подобрки литературы на сайте "Проблемы эволюции": Проблема эволюционных новообразований, http://evolbiol.ru/news.htm Причины прогресса, http://evolbiol.ru/progress.htm А. П. РАСНИЦЫН. Процесс эволюции и методология систематики. 2002. // Труды Русского энтомологического общества. Т. 73. С. 1-108. http://www.palaeoentomolog.ru/Publ/Rasn/methodology.html И. И. ШМАЛЬГАУЗЕН. Факторы эволюции. Теория стабилизирующего отбора. 1968. 450 с. http://evolbiol.ru/factory.htm И. И. ШМАЛЬГАУЗЕН. Организм как целое в индивидуальном и историческом развитии. 1982. http://rogov.zwz.ru/Macroevolution/org_kak_tsel.doc Б. М. МЕДНИКОВ. Аналогия (параллели между биологической и культурной эволюцией). 2004 // Человек. № 1-4. http://vivovoco.rsl.ru/vv/papers/men/medn/analogy.htm |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||