|
||||
|
|
Нейрофизиологическая база представлений К. ЛоренцаСледующий шаг, который сделал Лоренц и развили его последователи, — это попытка соотнесения гипотетической «специфической энергии действия» с имевшимися в тот период данными о реально происходящих в нервной системе процессах. Большую роль сыграли при этом данные о наличии автоматических спонтанных процессов в центральной нервной системе (цнс). В тот же период, когда Лоренц (1937), исходя из наблюдений за поведением животных, постулировал представление о внутренних факторах, определяющих характер их проявления, немецкий физиолог Э. фон Хольст собрал и обобщил факты, которые стали нейрофизиологической базой концепции Лоренца. Первоначально этологи пытались использовать для объяснения получаемых данных рефлекторную теорию, однако постепенно накапливалось все больше фактов, которые не могли быть объяснены с ее помощью. В соответствии с классической рефлекторной теорией всякое поведение является реакцией на внешний или внутренний стимул. Афферентный нерв проводит возникающее в рецепторе возбуждение в спинной мозг, в сером веществе которого находится соответствующий «рефлекторный центр». Далее возбуждение передается на эфферентный нерв. Эти три элемента составляют так называемую «рефлекторную дугу». Более сложные цепи поведения — это цепи отдельных рефлексов. Согласно этой теории, например, у угря при плавании сокращение и расслабление одного мышечного сегмента туловища механически возбуждает проприрецепторы следующего сегмента, а они в свою очередь возбуждают его рефлекторную дугу, что вызывает сокращение мышц этого сегмента. Однако фон Хольст, изучавший плавание угря обнаружил, что эта гипотеза не выдержала экспериментальной проверки. До фон Хольста наиболее распространенным являлся следующий экспериментальный метод: искусственно деафферентировали цнс, а затем путем периферической стимуляции вызывали периферическую реакцию, которая и доказывала, что цнс функционирует как рефлекторный механизм. Фон Хольст изменил эту методику и сосредоточил внимание на активности самой цнс. Он показал, что в изолированной брюшной нервной цепочке земляного червя наблюдаются разряды ритмических импульсов, точно соответствующие сокращению сегментов ползущего червя. В дальнейшем, исследуя механизм плавания у угря, фон Хольст прибег к этому же методу. Если фиксировать средние сегменты тела угря, не давал им сокращаться, то, согласно рефлекторной теории, задние сегменты не должны придти в движение, не получив раздражения от соседних сегментов. Однако на самом деле через промежуток времени, соответствующий несостоявшемуся сокращению фиксированных экспериментальных сегментов, задние сегменты тела также приходят в движение. Это может быть показано еще более четко, если перерезать дорсальные корешки спинного мозга, передающие сенсорные импульсы. Оказалось, что несмотря на деафферентацию угорь сохраняет способность к хорошо координированным плавательным движениям. Это должно означать, что все плавательные движения полностью запрограммированы в цнс и выполняются независимо от внешних раздражений, а не по механизму рефлекторной дуги. Существовал и ряд данных, которые лишь с натяжкой укладывались в рамки рефлекторных представлений. Было обнаружено, например, что у децеребрированных кошек две полностью деафферентированные антагонистические ножные мышцы ритмически сокращаются. Это привело к выводу, что в цнс существует эндогенный автоматизм, не зависящий от внешних раздражений. Позднее Вейсс пересадил участок спинного мозга и зачаток конечности у зародыша аксолотля. Развивающийся зачаток иннервировался пересаженным эмбриональным спинным мозгом. Поскольку мотонейроны развиваются быстрее сенсорных нейронов, их отростки, прорастая, достигают пересаженной конечности раньше, чем отростки сенсорных клеток. В тот момент, когда произошло афферентное соединение, конечность начала двигаться. Хотя совершаемые ею движения и не были полностью характерными для ходьбы, тем не менее в них можно было обнаружить примитивные формы координации, включая переменное сокращение мышц антагонистов. Этот факт также демонстрирует наличие эндогенного автоматизма в цнс и существование морфологических структур, обеспечивающих его проявление. Аналогичные явления были продемонстрированы и на других позвоночных. Деафферентированные головастики, жабы и рыбы сохраняли способность плавать, нормально координируя движения, до тех пор, пока у них оставался неразрушенным хотя бы один чувствительный нерв. Следовательно, для поддержания плавательных движений необходим минимальный уровень афферентной импульсации, которая способствует поддержанию уровня возбуждения в соответствующих нервных центрах (или «специфической энергии действия» по Лоренцу). Большое значение в этом плане имели работы Рёдера, который и обнаружил недостаточность рефлекторной теории для объяснения некоторых сторон поведения насекомых. В последующие десятилетия эти данные о возможности спонтанной активности в цнс насекомых получили широкое экспериментальное подтверждение. По представлениям Рёдера, нервные элементы, лежащие в основе спонтанного и рефлекторного поведения, различаются, в частности, по порогу возбудимости. Порог раздражения первых может быть настолько низким, что инстинктивная реакция способна осуществляться помимо видимых внешних раздражителей. Влияние уровня возбуждения в соответствующих нервных центрах на характер протекания инстинктивных реакций было подтверждено работами немецких ученых Э. фон Хольста и У. Сент-Поль (1960). В зависимости от локализации раздражаемой током структуры ствола мозга кур они наблюдали либо элементарные поведенческие реакции (клевание, повороты головы), либо сложные акты поведения (ухаживание за отсутствующими самками). Сложные реакции протекали менее константно, чем простые. Они зависели от состояния животного, от предшествующих стимулов от параметров тока и всей совокупности действующих на птицу стимулов. Например, при слабом токе петух бросался на чучело хорька, на которое до этого не обращал внимания, а при усилении тока такая же реакция проявлялась на неспецифический раздражитель — кулак. Гидравлическая модель.Лоренц предложил гипотетическую модель осуществления реакций типа завершающих актов, общие принципы которой он заимствовал из гидравлики. В настоящее время гидравлическую модель можно считать глубоким анахронизмом в этологии, представляющим лишь исторический интерес. Ввиду этого мы остановимся на ней очень кратко (рис.1). Накопление «специфической энергии действия» представлено в модели в виде постепенного накопления воды в резервуаре. 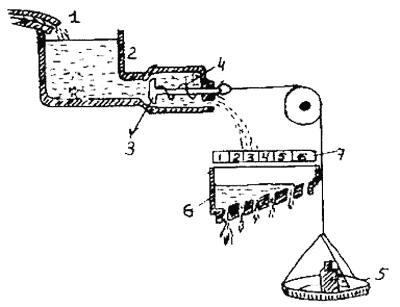 Рис. 1. Схема гидравлической модели для объяснения регуляции поведения (см. Меннинг, 1982) 1 — кран, 2 — резервуар, 3 — клапан, 4 — пружина, 5 — чашка весов, 6 — лоток 7 — шкала Давление жидкости внутри него все время повышается. Отток жидкости из резервуара (аналогизируемый с активностью животного) возможен по трубам, которые блокируются клапаном. Этот клапан может открыться при достаточно большом давлении (накоплении жидкости в резервуаре), а также при воздействии извне — при действии тяжести гири, приложенной к клапану. Такое внешнее воздействие аналогизируется с влиянием внешних раздражителей, специфичных для данного поведенческого акта. Нарастающее давление и действие внешней силы направлены одинаково и могут суммироваться. Чем больше жидкости набралось в резервуаре, тем меньше усилий нужно приложить извне, чтобы открыть клапан, а иногда клапан может открыться и без внешнего воздействия. Этот момент моделирует появление активности «вхолостую», возникающей без видимых внешних сигналов. Эта аналогия может быть продолжена. Если клапан открылся лишь немного, жидкость будет вытекать слабо и попадет только в область нижнего отверстия специального наклонного лотка. Если же клапан откроется сильнее — жидкость потечет сильнее и попадет и на следующие отверстия. Этот лоток с отверстиями аналогизируется с разными типами двигательной активности, относящейся к данному поведенческому акту. Самое нижнее отверстие соответствует активности с наиболее низким порогом — например, некоторым проявлением поискового поведения. Другие отверстия соответствуют иным формам активности, обладающим более высоким порогом. Если резервуар опорожняется, т. е. иссякает; «специфическая энергия действия», — данное поведение больше продолжаться не может. Современная оценка гидравлической модели Лоренца.В определенных пределах предложенная Лоренцем модель хорошо описывает феноменологию инстинктивных действий, а также циклические изменения, происходящие в реактивности нервной системы животного к внешним раздражителям — снижение порога совершения действия, если оно долго не выполнялось, восстановление готовности к инстинктивным действиям после перерыва и возможность появления реакций на неспецифические раздражители. В течение некоторого времени она являлась стимулом к развитию новых исследований. Однако наряду с этим модель Лоренца неоднократно подвергалась критике, отчасти необоснованной, но отчасти и справедливой (например, механистичность построения). При создании модели принимаются в расчет лишь самые общие черты, отражающие схему работы моделируемой системы, а также ее соответствие известным из опыта фактам. Другое дело, что предложенная Лоренцем модель, хорошо объясняя очень многие из известных фактов, объяснить другие была не в состоянии. Однако это свидетельствовало лишь о том, что представление о запасании «специфической энергии действия» имеет ограниченное применение, и соответствующие термины и понятия не универсальны. Анализ применимости модели Лоренца к описанию поведения в сопоставлении с другой моделью, предложенной Дейчем, можно найти в книге Меннинга (1982). Плодотворность применения модели Лоренца мы иллюстрируем в гл.5 на примере анализа поведения кошек, выполненного Лейхаузеном. Современная оценка концепции Лоренца о врожденном разрешающем механизме опирается на большое количество экспериментальных фактов, полученных в последние годы. Подробный обзор современных взглядов по этим вопросам приводится в книге Меннинга. Очевидно, что и «специфическая энергия действия» и ключевые стимулы — это понятия, которые в переводе на язык современной нейрофизиологии выражаются такими терминами, как «специфическое побуждение», активация той или иной мотивационной системы, а также видоспецифическая избирательность перцепторного аппарата. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Вверх |
||||
|
|
||||